

Original paper: Spreading dynamics and wetting transition of cellular aggregates
In episode one of this series, I presented a research paper by Stéphane Douezan and his colleagues in which they studied a ball of cells (called a cellular aggregate) sitting on a flat surface. After introducing the concept of cellular aggregate wetting by comparing it to the classical system of a drop of water, today I present the main part of the paper which looks at the dynamics of spreading of the cellular aggregate. I strongly suggest that the reader reads the first post before reading this one.
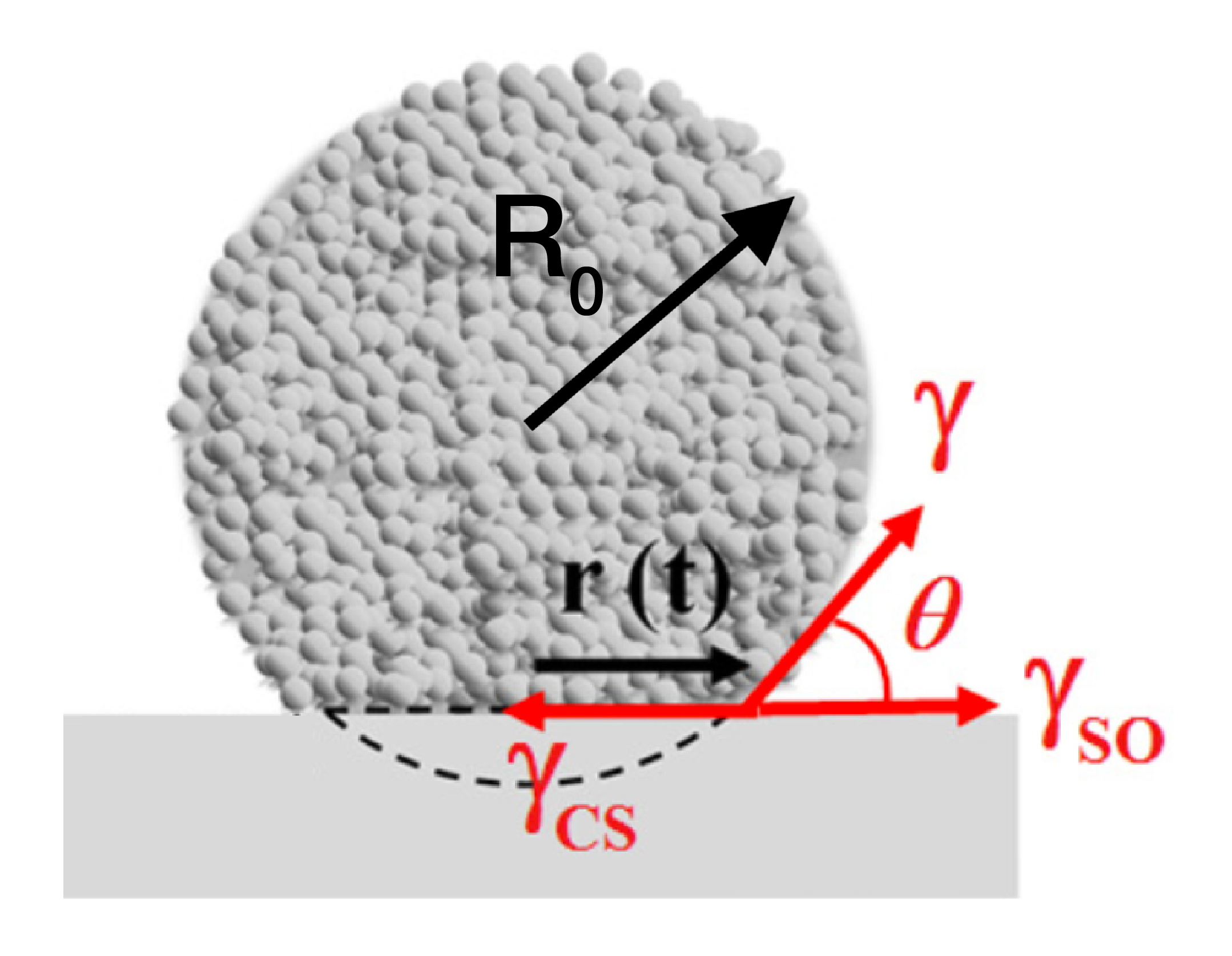
As introduced previously, the spreading of a cellular aggregate is set by the surface tensions of the three interfaces: cells-substrate ($latex \gamma_{CS}$), cells-medium ($latex \gamma$), substrate-medium ($latex \gamma_{SO}$). The spreading can be controlled by finely tuning two adhesion energies: the cell-cell adhesion ($latex W_{CC}$) and the cell-substrate adhesion ($latex W_{CS}$) [1]. The authors of this paper set $latex W_{CC}$ by controlling the level of E-cadherin (a molecular glue between cells), and $latex W_{CS}$ by varying the concentration of fibronectin (a molecular glue between the cells and the substrate) deposited onto the substrate.
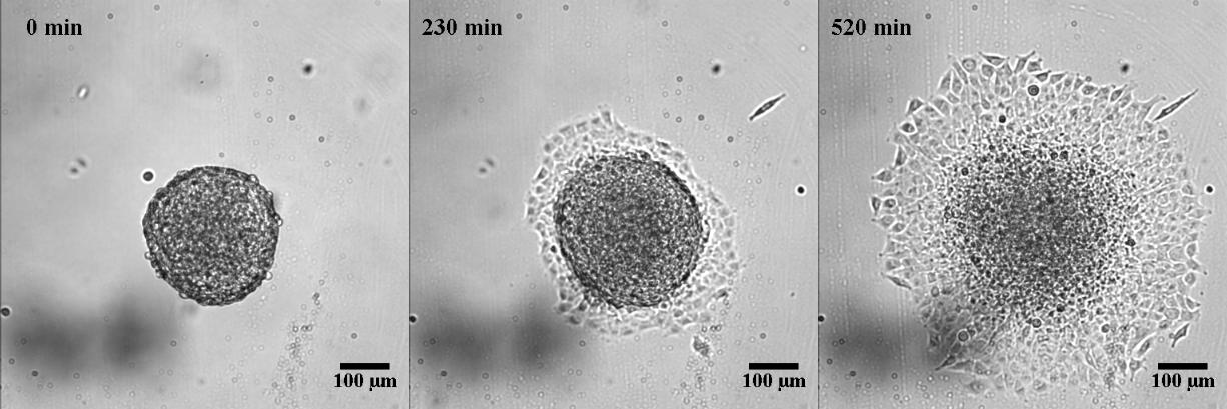
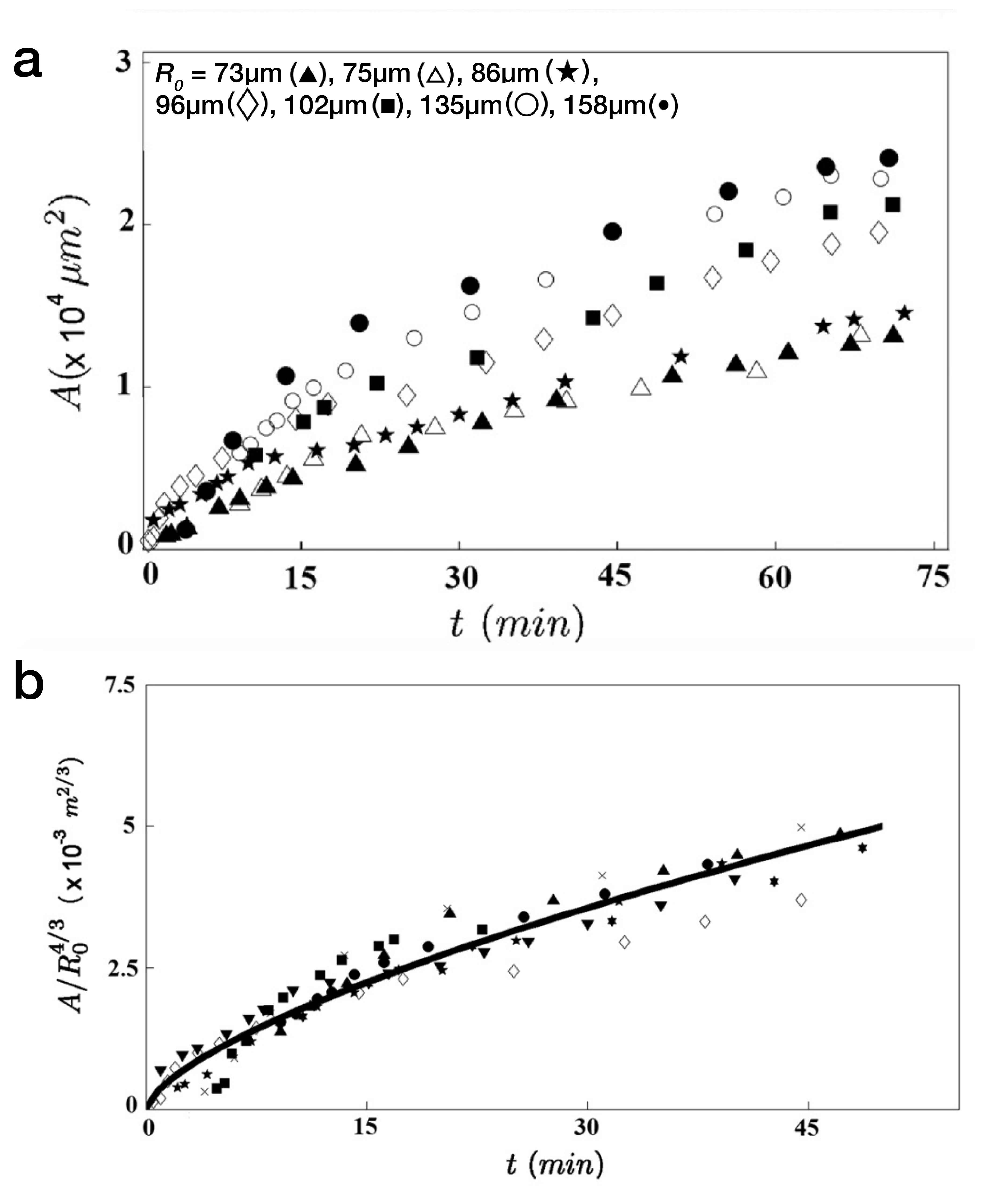
To characterize the dynamics of spreading, Stéphane Douezan and his colleagues measured the area of the cellular aggregate in contact with the surface with respect to time. The authors noticed two distinct regimes: at short times (the first hour) the cellular aggregate flattens, and at longer times, it forms a film which spreads completely. In the first regime, they observed a non-constant spreading speed. More interestingly, it depends on the cellular aggregate size: the bigger the aggregate, the faster the spreading (see Figure 2a).
To understand this non-trivial spreading dynamics, the authors investigated in detail what is driving and what is preventing the cellular aggregate from flattening at short times. The contact area expands because the adhesion between the cells and the substrate is more favorable than the cell-cell adhesion. So increasing the cell/substrate adhesion $latex W_{CS}$ should increase the speed of spreading. On the other hand, the process is slowed down by the friction of the cells: there is a so-called viscous dissipation, like when you pour honey, the more viscous the honey the longer it takes to flow. So increasing the viscosity, should decrease the speed of spreading. The authors expressed the energy of these two antagonist contributions to the speed.
First, the energy gain is the work per unit of time of the capillary force $latex F_c$ [2]:
Energy gain = $latex 2\pi r F_c \frac{dr}{dt}$ [3]
At early times, the contact angle is very small, so the capillary force $latex F_c$ can be simplified: $latex F_c = W_{CS} + \gamma (cos \theta -1) \approx W_{CS}$ . In this way, $latex F_c$ can be replaced by the constant $latex W_{CS}$ in the expression of the energy gain.
Second, the authors show the dissipation is expressed by $latex \eta (\frac{dr}{dt})^2 \frac{r^3}{R_0^2} $ where $latex \eta$ is the cellular aggregate viscosity. Per conservation of energy, the energy gain should be exactly compensated by the viscous dissipation. Thus, by equating these two energies and integrating $latex r$ over time, we have the time variation of $latex r^2$ that follows a power law [4]:
$latex r^2 \propto R_0^{4/3}\frac{W_{CS}}{\eta}^{2/3} t^{2/3}$, with $latex R_0$ being the aggregate initial radius.
This dynamics of $latex r^2$, which is proportional to the contact area, indeed depends on the aggregate size $latex R_0$ in a consistent manner with the experimental observations: the bigger the aggregate, the quicker it spreads.
So if the law is valid, rescaling the measured area by $latex R_0^{4/3}$ should remove the dependency on the size of the cellular aggregate. This is exactly what they saw: all the data points collapsed on the same curve (Figure 2b). There is something even more interesting here: fitting the spreading curve gives an estimate of the ratio $latex W_{CS}/\eta$, two variables which are difficult to measure. Of course, this is only a ratio, which does not provide absolute values for these two variables but it possible to measure relative changes by playing with some biological parameters. For instance, as mentioned above, the authors can tune the cell-cell adhesion energy using genetic tools (see the first post to understand how they measure it) and the cell-substrate adhesion by coating the substrate with different concentrations of an adhesive molecule. In this way, they quantitatively described how the viscosity decreases when the intercellular glue expression — the E-cadherin — is reduced, see Table 1. Similarly, they studied the relative change of the cell-substrate adhesion energy depending on the substrate coating.
Table 1. Relative change of the viscosity depending on the E-cadherin expression.
| E-cadherin level (controls cell-cell adhesion energy) | 21% | 48% |
| Relative viscosity to the 100% expression | 42% | 57% |
To summarize, Stéphane Douezan and his colleagues were able to explain what is driving the initial flattening of the aggregate at short times by showing how this dynamics depends on the aggregate size, and they were able to estimate the ratio of the cell-substrate adhesion energy over the viscosity.
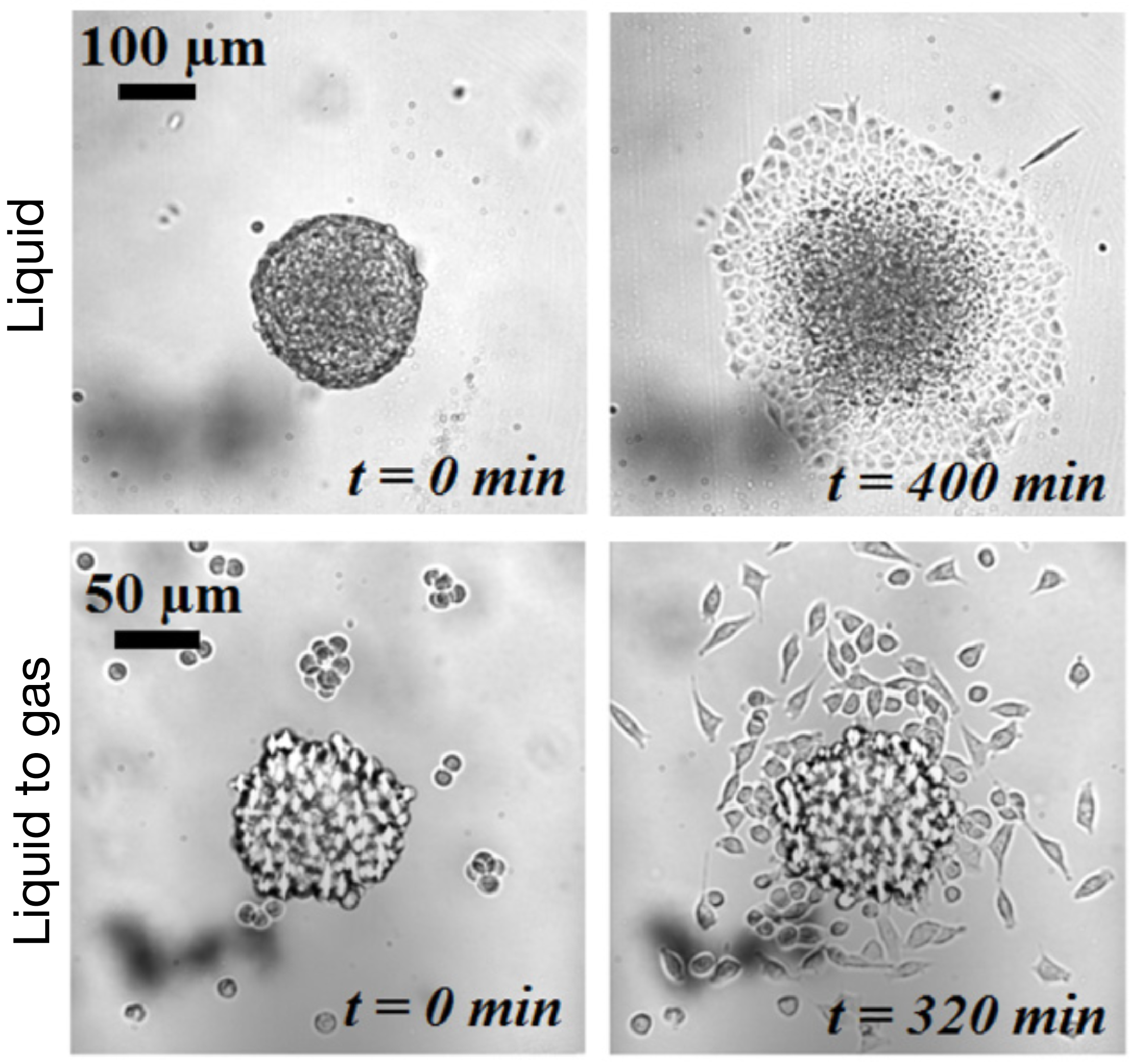
After studying this short-time regime, the authors analyzed the spreading at longer times. Depending on the cell-cell adhesion energy, they noticed two behaviors: either the aggregate flows as a cohesive two-dimensional sheet of cells (like a liquid) when the adhesion is strong, or individual cells escape from the aggregate (analogous to a liquid-to-gas transition) when the adhesion is weak. These two behaviors are shown in Figure 3 and in movie 2 and 3 of the supplementary data. This phenomenon could be used to model an invading tumor for which the biological parameters that control the transition between two kinds of spreadings can be precisely tuned.
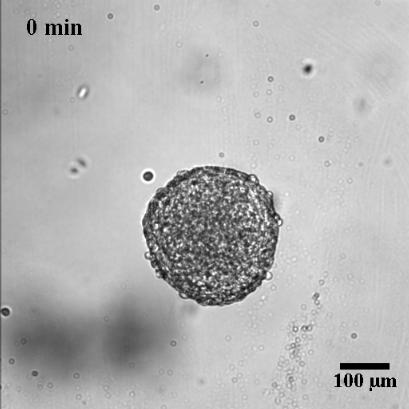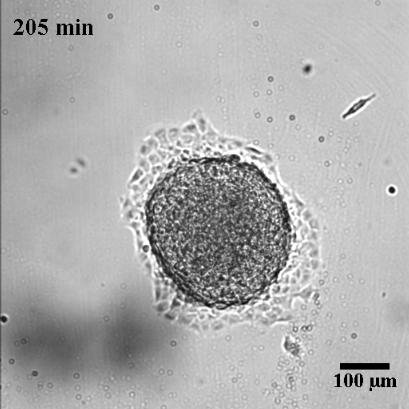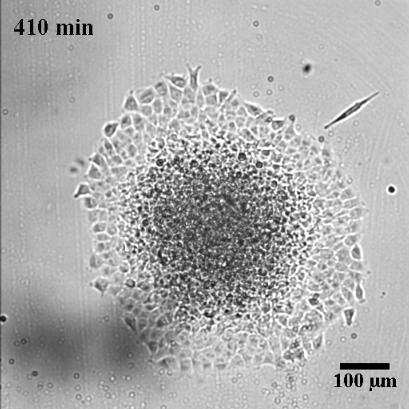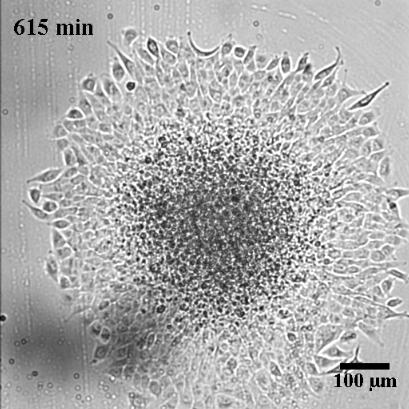
In this paper, the authors successfully captured the complexity of a biological system with a predictive law of spreading. By measuring well defined physical variables, such as the viscosity and the cell-substrate adhesion energy, they were able to quantify how cells bind to each other or to their environment. These complex biological processes, which involve many different molecular actors, are often described in a qualitative way. Even more interestingly, the authors showed how they could tune these physical variables by controlling some biological parameters, which directly shows their implications in the processes mentioned above. The approach taken in this paper is very elegant as biology often fails to be predictive because of the important complexity of the processes at stake.
[1] The adhesion energy of an interface is the work that should be spent by unit of area if one were to break this interface. The stronger the energy, the more stable the adhesion. Therefore like the surface tension, it is an energy density (unit: $latex J/m^2$). As a reminder from the previous post, the two adhesion energies can be expressed by the surface tensions: $latex W_{CC} = 2 \gamma$ and $latex W_{CS} = \gamma_{SO} + \gamma – \gamma_{CS}$.
[2] The capillary force is the sum of the components of the three tensions along the tangent axis to the substrate. This force per unit of length is basically the force that pulls on the drop: $latex F_c = \gamma_{SO} + \gamma cos(\theta) – \gamma_{CS}$.
[3] The differential expression of the energy gain is obtained through the following reasoning: during an infinitesimal duration of spreading $latex \delta t$, the radius of the contact line increases by $latex \delta r$. So the infinitesimal work of the driving force $latex F_c$ is: $latex \delta W = perimeter * F_c * \delta r = 2\pi r F_c \delta r$.
[4] A power law is simply a mapping of a variable at some power. They are usually presented on log-log plots, as they appear as a straight line, for which the slope is the power of the function.
Disclosure: The second author of this paper is my Ph.D. supervisor. However, she did this work while she was a postdoc. Consequently, I have never been involved in this work.
One Reply to “The living silly putty, episode 2: the spreading!”