

Original paper: Spreading dynamics and wetting transition of cellular aggregates
Disclosure: The second author of this paper is my Ph.D. supervisor. However, she did this work while she was a postdoc. Consequently, I have never been involved in this work.
Have you ever noticed how drops of water have different shapes on a clean piece of glass and in a frying pan? The frying pan surface is coated with a hydrophobic (“water-repellant”) molecule so it does not stick to food, which typically contains a lot of water. As a result, a drop of water will take a roughly spherical shape to reduce as much as possible its area of contact with the frying pan. If a surface has an even more hydrophobic coating than a frying pan, the drop can even reach a perfectly spherical shape (this is called ultrahydrophobicity, or the lotus effect). At the opposite extreme, glass is said to be hydrophilic (“water-loving”) — when placed on a clean piece of glass, a drop of water tries to increase its surface of contact much more than a droplet on a hydrophobic frying pan. Depending on the hydrophilicity of the underlying surface — which is known as the substrate — the drop has a well-defined area of contact. The interaction between fluid interfaces and the solid surfaces is a very well studied field of soft matter called wetting. Researchers in this field investigate how the three different interfacial energies — interfaces between water and substrate, between water and air, and between substrate and air — dictate what shape a droplet takes, and how it spreads across the surface.

{kind=link}
Today’s post is the first one of a series of two (click here for the second one), which deals with the work of scientists who replaced the drop of water by balls of living cells called cellular aggregates. They deposited these aggregates onto different surfaces to carry out an experiment analogous to the spreading of water droplets. In the case of a drop of water, only the physical and chemical interactions between molecules determine the shape of the drop. When a drop sits on a substrate there is an interface between the water and the substrate. If the chemical interactions between the substrate and the water are not favorable (hydrophobic), the price to pay will be a large interfacial energy. As every system in physics, it tries to reduce its overall energy by reducing the area of contact. The drop shrinks, like the ones you can see in your frying pan. But as it shrinks, the interface area between the substrate and air increases by freeing the surface. And, as the volume of water is fixed (we consider a no-evaporation situation), the surface of the drop in contact with air changes too. Therefore, the drop shrinks or spreads up to a point for which the sum of the three interfacial energies is minimized.
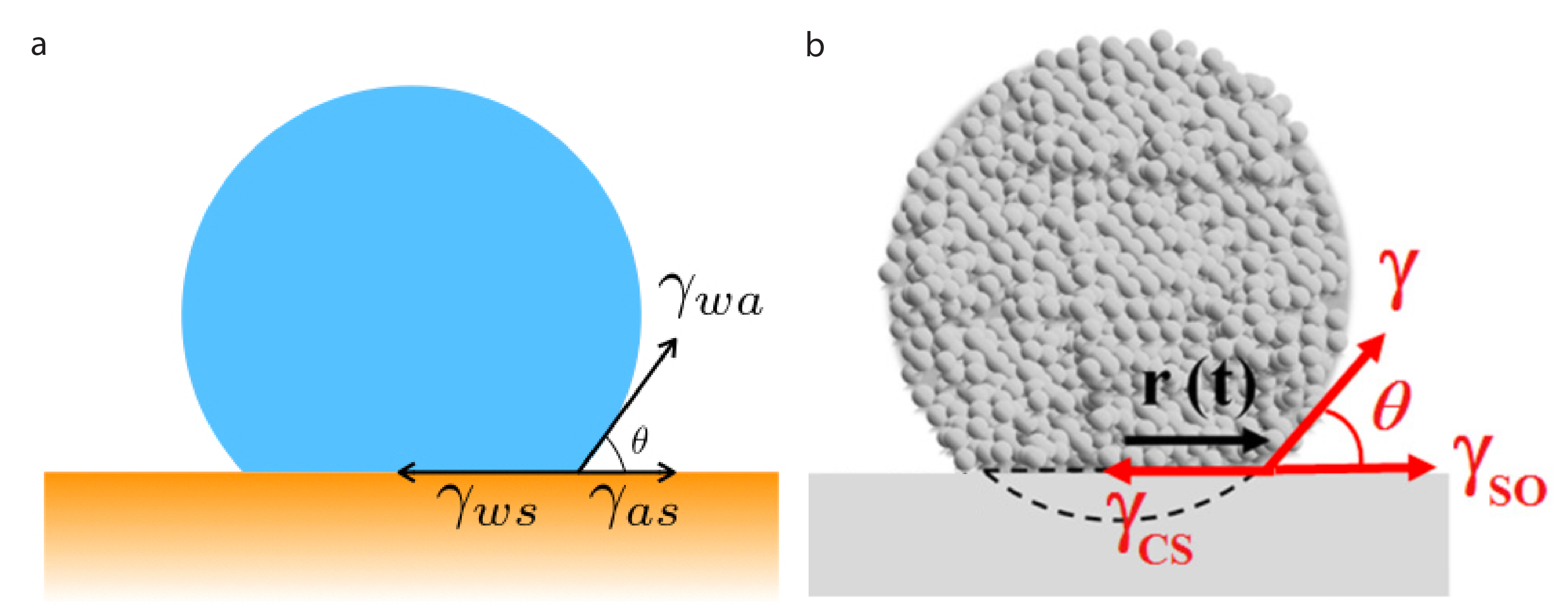
The shape of the drop can be described by the contact angle ?, which can be used to predict the interfacial tensions, which quantify how the interfacial energies change when the areas of contact are changed. A tension is a force divided by a length. So, we can write the equilibrium of tensions on the point where the three interfaces meet (1):
$latex \gamma_{ws} = \gamma_{as} + \gamma_{wa} cos\theta$
To predict if the drop will spread on the surface the wetting coefficient $latex S$ can be defined as:
$latex S = \gamma_{as} – ( \gamma_{wa} + \gamma_{ws} ) $
$latex S = – \gamma_{wa} (cos\theta -1)$ (by using the equilibrium of tensions) (2)
This expression shows that there is a partial wetting ($latex \theta$ between $latex 0$ and $latex \pi$) if $latex S<0$. If $latex S>0$, the drop spreads completely: that is, the droplet covers the substrate with an infinitely thin fluid film.
Now, what happens if we consider a ball of cells instead of a drop of water? Since the pioneering work of Malcolm Steinberg, we know that cellular aggregates can behave as liquids over long times. If you were to poke a piece of biological tissue, it would resist at short times (less than a dozen of seconds to a couple of minutes) but on the long run, it would start to flow. As every liquid, a surface tension builds at its interface. For instance, a rough ball of cells in suspension will round up over time to minimize its ratio area/volume. As presented in a previous post, the surface tension is a physical value that can also be defined for biological tissues even though its nature is very different from the one of purely physical systems.
To further investigate the role of surface tension in living tissues, Stéphane Douezan and his colleagues decided to study how the biological properties of these aggregates of cells can influence their liquid behavior. The first property they considered was the “stickiness” of the cells, also known as cell-cell adhesion. Cells produce a large number of molecules at their surface which allows them to sense and interact with their environment. E-cadherin is an important molecule that acts like a kind of glue between cells, allowing them to stick to their neighbors. Using genetic tools, the researchers grew cells with different levels of E-cadherin, making them more or less sticky with respect to the others. By using micropipettes to pull on two sticking cells until they broke apart, the researchers then measured the energy of cell-cell adhesion. Integrating the force exerted during the separation and dividing by the contact area leads to the cell-cell adhesion energy $latex W_{CC} $.
The authors played with a second property too. In living tissues, cells interact with the extracellular matrix — a scaffold of molecules that gives the tissue its structure. One of the important molecules of the extracellular matrix is called fibronectin. By coating the glass substrate with different concentrations of fibronectin, the researchers could finely tune the adhesion of the cells to the substrate. To measure this adhesion: the researchers define the cell-substrate adhesion energy, $latex W_{CS} $.
In order to know if an aggregate will spread, the wetting coefficient $latex S = \gamma_{SO} – (\gamma + \gamma_{CS})$ must be evaluated. However, not all these tensions can be measured directly, so they must be expressed in term of the energies we can measure $latex W_{CS}$ and $latex W_{CC}$. A classical approach to connect the adhesion energy to the tensions is to write the balance of tensions if we were to break an interface. For instance, to separate a cell-cell interface, two new interfaces (between the cells and the surrounding fluid) must be created so, by energy conservation: $latex W_{CC} = 2 \gamma$. Similarly, breaking a cell-substrate interface requires creating an interface between the cell and the surrounding fluid, an interface between the substrate and the fluid, and removing a cell-substrate interface, so: $latex W_{CS} = \gamma_{SO} + \gamma – \gamma_{CS}$.
Therefore, the wetting coefficient becomes $latex S = W_{CS} – W_{CC}$. If $latex S>0$, the energy of adhesion with the substrate is larger than the cell-cell adhesion energy, and the aggregate spreads completely. In this case, the dynamics of spreading can be monitored, as you can see in this video of wetting (video S1).
In the next post, I will present the dynamics of spreading, where the cellular aggregate literally behaves as a chunk of silly putty!
(1) In reality, the three interfaces meet at the line that circles the drop, but since the system has a circular symmetry, it makes more sense to write the tension balance on a point instead of writing the force balance all along the circle.
(2) Usually, $latex \theta$ is defined as the complementary angle ($latex \theta ‘ = \pi /2 – \theta$), so $latex S=( cos\theta ‘ – 1 ) \gamma_{wa}$ . But here I decided to use the same definition as the authors for the sake of consistency.
One Reply to “The living silly putty”