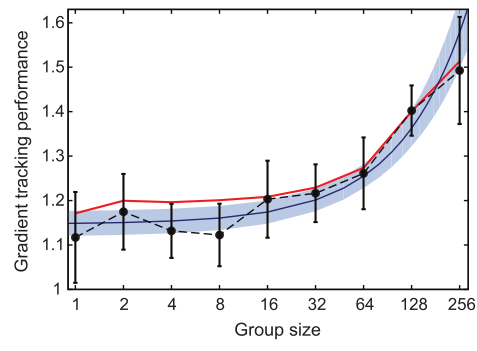
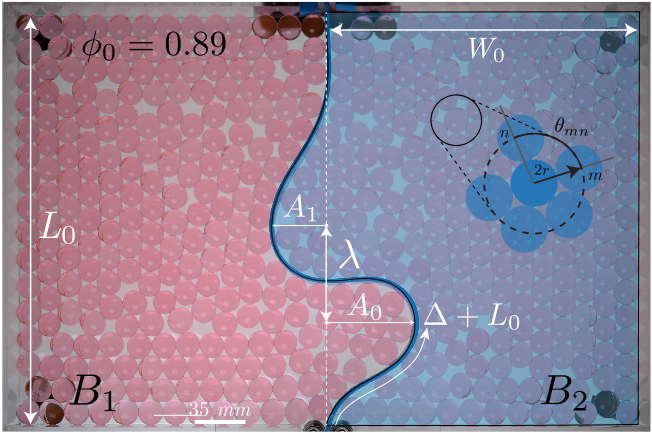
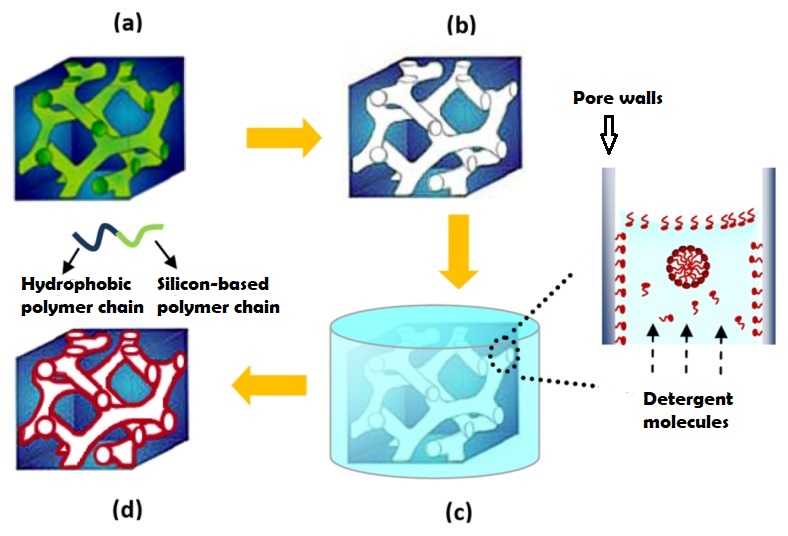
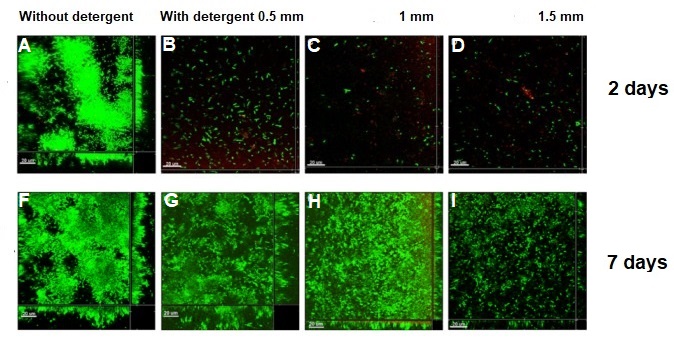
“ESOF (EuroScience Open Forum) is the largest interdisciplinary science meeting in Europe. It is dedicated to scientific research and innovation and offers a unique framework for interaction and debate for scientists, innovators, policy makers, business people and the general public.” [1]
This year ESOF was in Toulouse, and I was fortunate enough to be able to attend, so I want to share a few snippets of my time there, and my main takeaways.
There was a huge range of different topics on their programme from science communication and careers, to atomic clocks and plastic pollution. I found choosing between parallel sessions was often difficult. But even though I went to a variety of different sessions, I am going to focus on one major theme kept cropping up. Namely, trust.
How can we, as scientists, build trust in science? How can we trust one another?
For this science and scientists need to be seen as credible. There are no quick fixes, but openness was touted by many at ESOF as a huge step towards building more trust. After all if ‘science is not finished until it’s communicated’ [2] then the public are a huge part of science! Not to mention that for most of us, we are in fact paid by taxpayers.
Here are some of the different types of openness that people discussed at ESOF2018:
- Open Access
Currently, many scientific journal articles are behind paywalls – this means institutions without access to a particular journal are locked out. Even when a journal allows institutes to post papers open access from the institute, this is often after long embargo periods restricting access to the latest science.
- Open Data
Even if a paper is open access. The methods and the data shown are often too little for experiments or analysis to be easily reproduced. For private and sensitive data – this is not possible but we can strive for the data to be as open as possible and only unavailable when necessary. If the data is accessible and readable, then anyone can reproduce their analysis, particularly if codes are also made accessible and usable.
- Open Communication
To share our research with the public, making something understandable to people outside your field is not enough. We need to open a dialogue between scientists and the public so that the outreach activities are catered to their interests. In particular for influencing policy, the people who our science affects need to be heard.
- Open Science
Science is a part of our culture, our history and our future. It would be difficult to find anyone who isn’t affected by science. Allowing people to take part is great for building trust. New initiatives and new technologies are opening the ‘ivory tower’ of academia. Citizen science or crowd-sourced science can bring together the public and scientists, for mass collection of data. And new make-spaces, fab-labs and open source software are making it easier for people to conduct their own experiments, build their own devices and explore science in their free time.
A bit of a culture shift needs to occur within science to highlight these aspects of science. I think, most scientists agree openness is important but often these ‘extra’ activities get put on the back burner as publications, teaching responsibilities and funding applications dominate our time. So, more time needs to be invested in openness, but not at the expense of an individual scientist’s career (or home life). Hiring practices, funders and fellow scientists need to reward and encourage openness within science. So I think it is great that so many people from a range of different places were talking about how to increase openness.
Overall I thought ESOF2018 was a very friendly conference, with so many passionate people working towards a better scientific process. To hear them talk so passionately, either in how science is funded, publishing, science policy, collaborations with industry, scientific careers, science communication and even the science of science communication was a fantastic experience.
Let us know if you have any experiences, thoughts, or difficulties on accessing science or how to open up science over twitter (@softbites17 or @emilyeriley) or in the comments down below.
[2] Quote from Sir Mark Walport, who was Chief Scientific Adviser to the UK government.