

Original Article: Bacteria exploit a polymorphic instability of the flagellar filament to escape from traps
No one likes being stuck. Whether you are in a car stranded in mud or stuck in a dead-end job, continuing normal behaviour is unlikely to help. Whereas we can see approaching hazards and dead-ends and try to avoid them, bacteria must blindly swim through passageways and channels that are of a similar size to themselves, often resulting in the cell becoming trapped. So, how does a bacterium change its behaviour to free itself?
In today’s paper, Kuhn and colleagues report a new type of bacterial motion used to escape from microscopic traps. Like the car stuck in slippery mud, forward-reverse strategies do not free the cell. The bacteria adopt a new swimming setup wherein their swimming appendage is wrapped around the cell body. This new coiled swimming was only discovered in 2017 and currently has only been observed for two bacteria species [1].
For bacteria, which are only a couple of micrometres, the viscosity of the swimming media dominates over any inertial effects. When we swim in water inertia dominates, so we can move to some extent with only one propelling action. However, when there is no inertia, if you stop propelling, you will immediately stop moving. To overcome the limits of their drag dominated environment, many bacteria use thin appendages called flagella.
Bacteria flagella are the only natural structures that generate propulsion via rotary motors. Much like a boat propeller, the bacterial flagellum has a rotary motor at its base, embedded in the cell body that rotates its propeller 100 times a second. However, the bacterial propeller is different from propellers we recognise. The bacteria flagella has two components: the flagellar filament and the hook. The flagellar filament is a helix, 1-3 times the length of the bacterium cell body, and the hook is a nanometre scale elastic segment connecting the filament to the rotary motor. Like a corkscrew, the rotation of the helix creates motion along the helix axis. Both the handedness of the helix, that is which direction the helix curls around its axis, and the direction of rotation determine whether a corkscrew goes forwards or backwards. Typically the bacterial flagellar filament is left-handed and rotates counterclockwise, looking from the end of the flagellum to the cell body, thus pushing the bacterium cell first at about 20 cell body lengths per second. If the same left-handed helix rotated clockwise, the bacterium would swim backwards – filament first – towing the cell body.
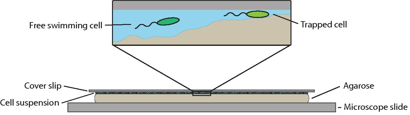
To recreate a bacterium’s natural environment, Kuhn and colleagues cover an uneven gel surface by a thin film of liquid with a thin glass slide above, as shown in Figure 1. The varying distance between the bumpy gel surface and the thin glass slide provided a micro-structured environment in which the bacteria could be observed. Their study focuses on a genetically modified soil bacterium that has a single flagellum placed at the end of a pill-shaped body a couple of micrometres in length. By watching the rotation and position of the fluorescently stained 20-nanometre-thick flagellar filament, they are able to determine when the bacterium is stuck and how structural changes enable its escape.
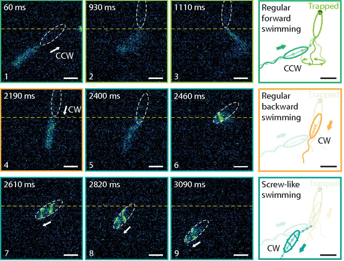
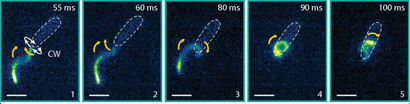
Compared to the free-swimming cell in a bulk solution, swimming close to surfaces increases the drag on the bacterium. The cell is trapped when the surfaces are close enough to increase the drag on the cell head above the thrust. To attempt to free themselves, the bacterium switches between clockwise and counterclockwise flagellar rotation – a forward-reverse strategy. This is unsuccessful. A successful escape only occurs when the flagellar filament wraps itself around the cell body, creating a conformation that has not been seen before. The wrapped filament is still helical, so rotation of the motor still creates propulsive forces along the helix axis. The cell then swims in a screw-like motion to release itself, as seen in Figure 2. It is yet unclear why exactly the coiled state enables the bacterium to escape. It could be the change in helical structure, the proximity of the flagellar filament to the cell body or the interaction of the flagellar filament with the nearby surface or even a combination of these fluid drag effects. Once the cell is free, the filament returns to its non-coiled state and normal swimming resumes. But how does this stiff inactive filament change its shape so drastically?
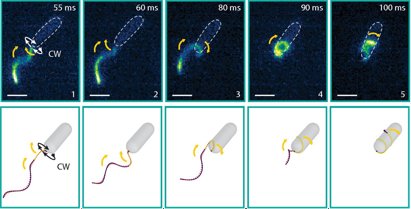
To induce the new swimming state in a bulk fluid environment, the researchers study the bacterium freely swimming in solutions of increasing viscosity. The increased viscosity increases the force on the flagellum and was observed to increase the likelihood of the screw-like motion. The increased forces on the flagellar filament thus seem to be responsible for the drastic change in state. Changes in the helical structure are well known for other flagellated bacteria changing the diameter and even the handedness of the helix, but never as extreme as to wrap the flagellum around the cell. These well-known flagellum changes are due to a change in the atomic structure of the flagellum, so rather than bend elastically, the filament changes shape sequentially along the flagellar filament. As the researchers’ bacterium’s filament is constructed similarly, they ascribe their two swimming types to two atomic configurations available to the flagellum: one normal and one coiled, as shown in Figure 3. Although the coiled swimmer is much slower than the uncoiled swimmer when freely swimming, the coiled state is able to free the bacterium from micro-structured traps, giving the bacterium a significant advantage.
Understanding this new type [1] of bacterial motion is critical to know how a bacterium survives in its natural habitat. For which habitats is the screw motion most common? Are there many more species that can coil to recoil? Where does this screw motion fail? What are the key features of the wrapped state that allow the bacterium to escape? Not only will studying these features help to understand populations of bacteria, for example how they spread and find better habitats, but it could also help in the design of micro-robots within complex environments such as for targeted drug delivery in the body.
[1] The same wrapped swimming configuration has also very recently been observed for a multi-flagellated bacterium, P. putida [you can see the paper here]. This bacterium has 5-7 flagella positioned at one end of the cell. Contrary to the results described above, the transition was observed for cells swimming in a bulk fluid environment. In the coiled state, all 5-7 flagella wrap around the cell. Through modelling the different swimming states of the cell, Hintsche and colleagues show an increased diffusion of populations that are able to transition to a coiled state.