Original paper: Adhesion of Chlamydomonas microalgae to surfaces is switchable by light
“One day it’s fine and next it’s…” red? Microscopic algae depend on photosynthesis, so they follow the light. Previous research has shown that their swimming is directed towards white light but not to red light. New work shows that light-activated stickiness allows microscopic algae to switch between different movement methods.
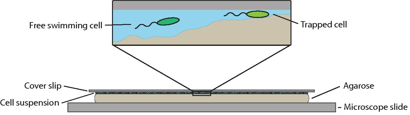
“This indecision’s buggin’ me” – should I stick or should I swim? Different types of motility are needed to move through different environments. Microscopic algae live in a variety of different conditions, including soils, rocks, and sands, all surrounded by water. In general, we can split these conditions into two groups: those where the algae move within the water, or those where the algae move across a surface. Today’s paper studies how a unicellular algae changes from its free swimming state to a surface attached gliding state.
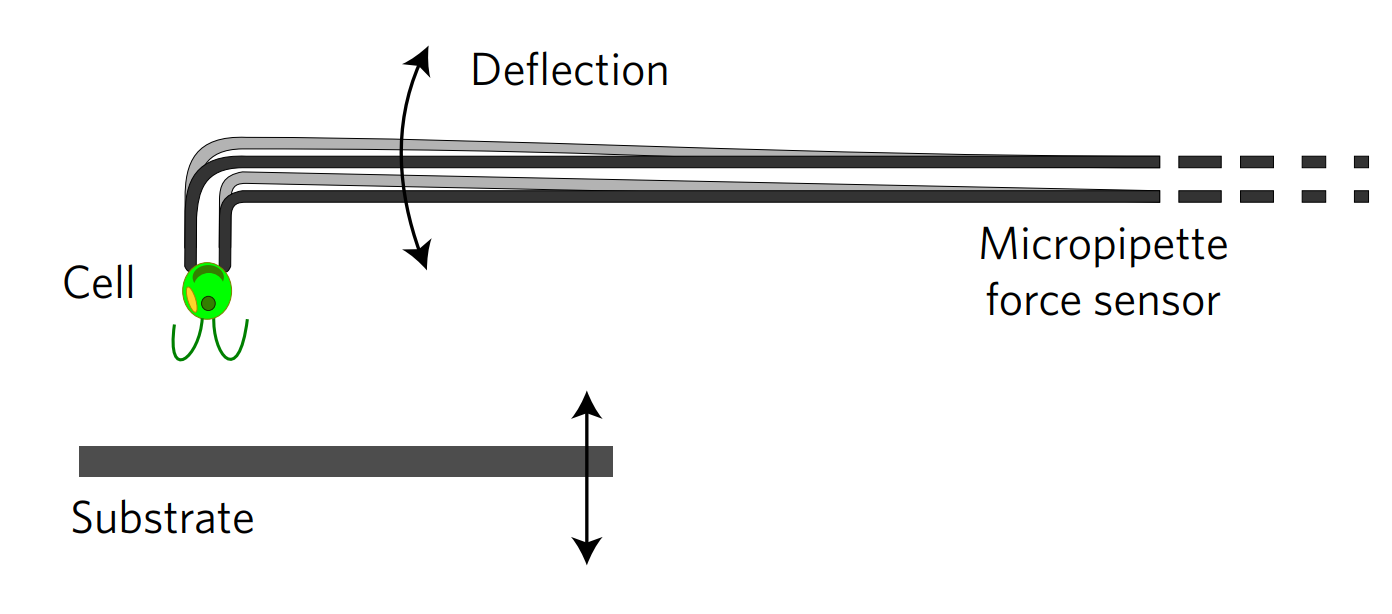
To transition between these two movement methods, the Chlamy must attach and detach from the surface. The researchers measure the force Chlamy exerts on a surface when it attaches using micropipette force microscopy, shown in Figure 2. This method uses a micropipette, which is a small glass tube, to hold a single Chlamy cell in place with suction. The surface is moved towards or away from the cell, deflecting the micropipette from its original position based on the force the cells exert on the surface. The relationship between deflection distance and force is measured beforehand with calibration experiments. So, during the experiment, micropipette deflection yields how strongly cells are stuck. To understand how this force relates to the two movements methods, let’s look at the results.
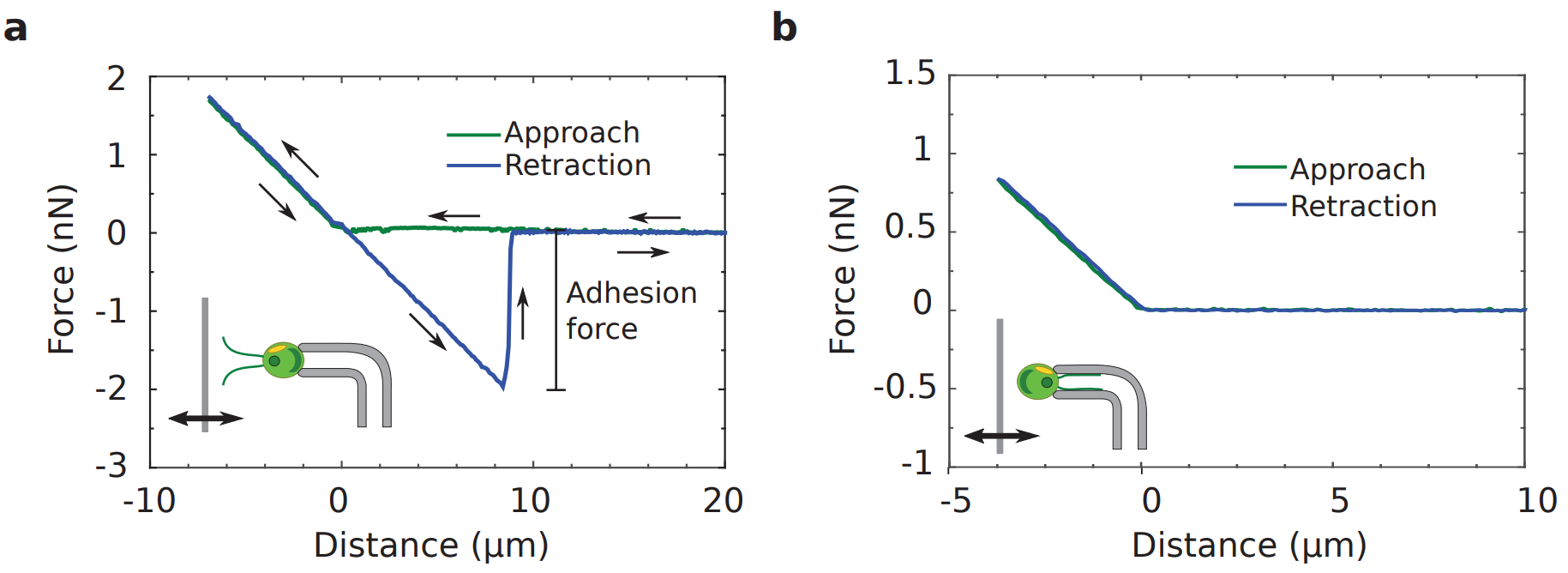
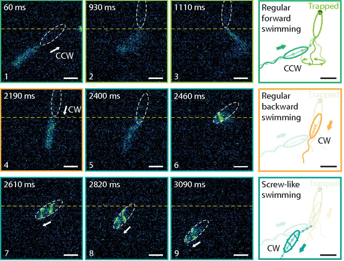
Figure 3 shows two force measurements, one where the flagella are facing the surface and another where the back of the cell is facing the surface. When the surface touches the flagella or back of the cell body, the micropipette is first deflected upwards, giving a positive force. As the surface is moved away, the micropipette moves back to its original zero-force position.
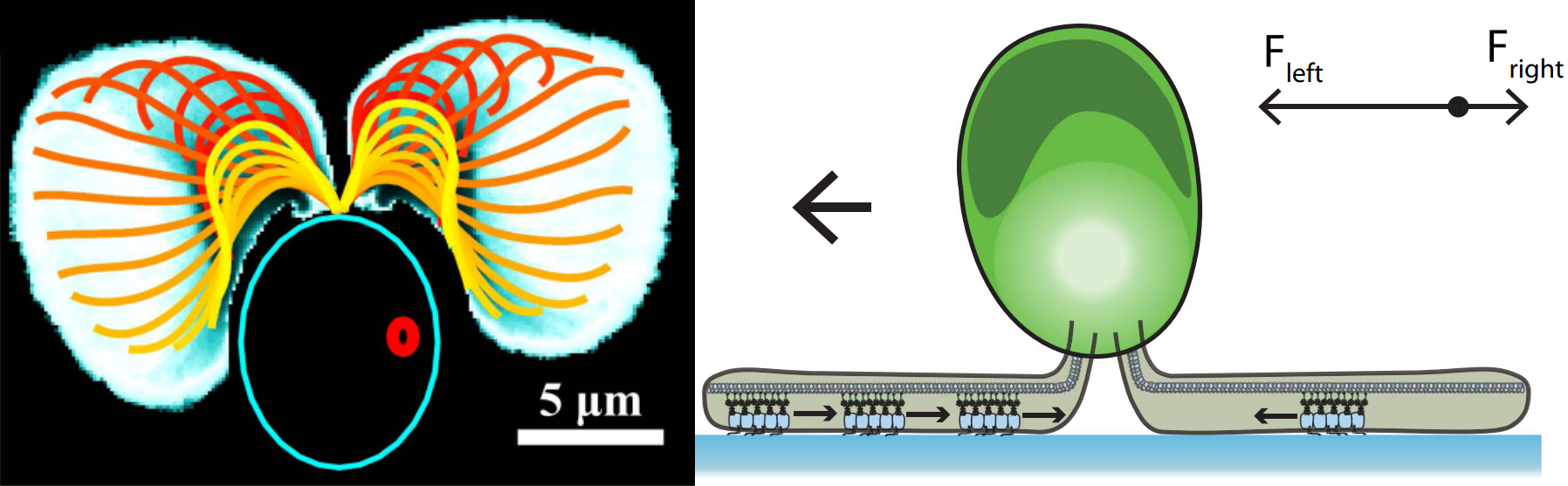
As the surface is moved further away, the flagella-facing cell and back-facing cell behave differently. The flagella-facing cell deflects the micropipette downwards, shown by the build-up of a largely negative force, whereas the back-facing cell does not deflect the micropipette and no force is exerted. This means that the flagella-facing cell sticks to the surface, whereas the back facing cell does not stick.
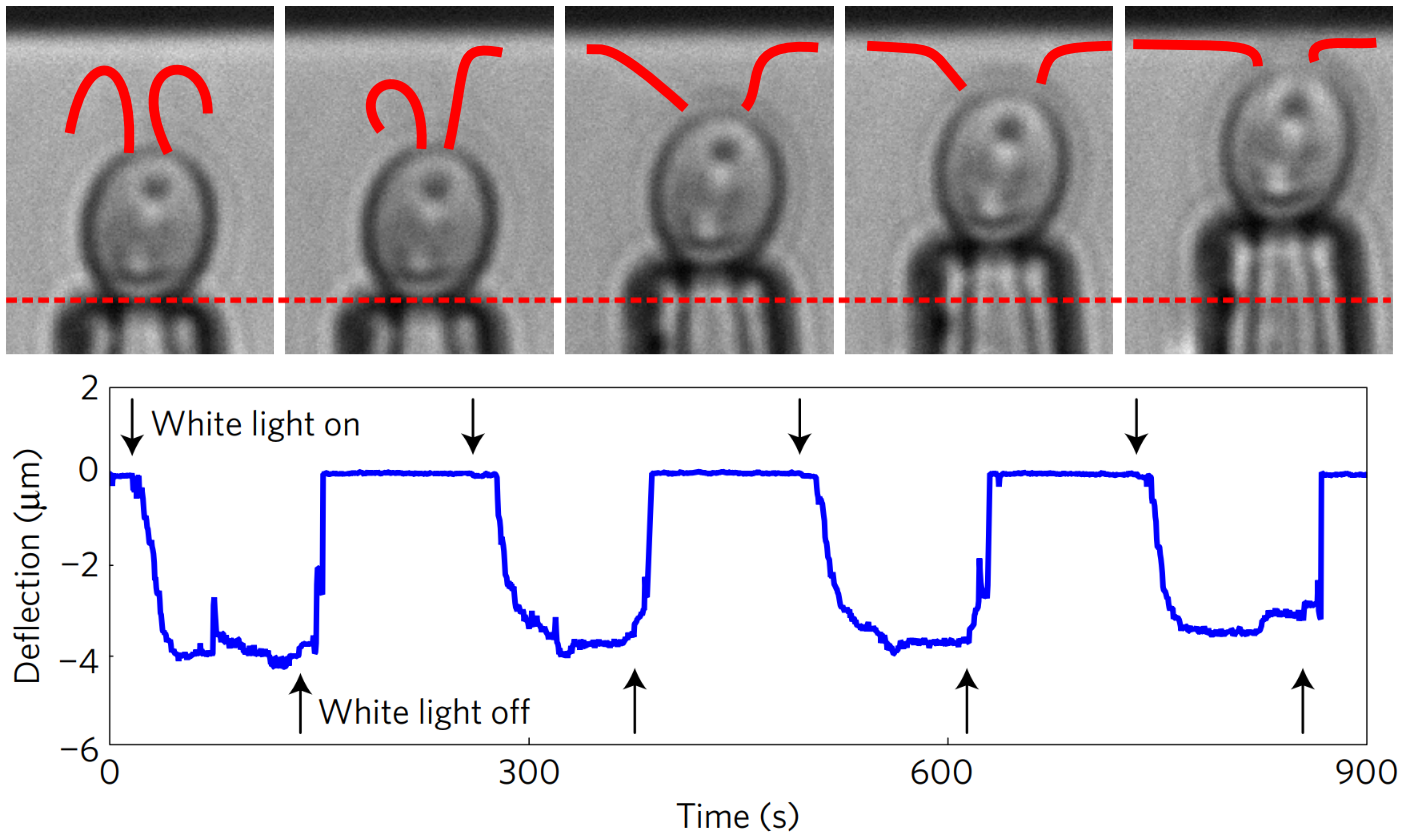
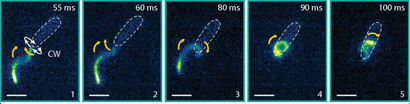
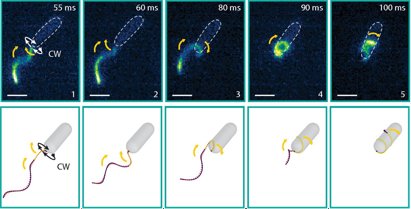
The flagella not only stick but actively pull themselves towards the surface. At the top of Figure 4, we see the flagella touch the surface during their swimming beat cycle. First, just a small part of one flagellum is stuck to the surface. Then, the flagella actively pull themselves towards the surface until both are completely stretched out and ready for gliding. This process is reversible: as the light is turned on and off, so is the adhesion force. The Chlamy can pull themselves up again and again – transitioning between their stuck and free state.
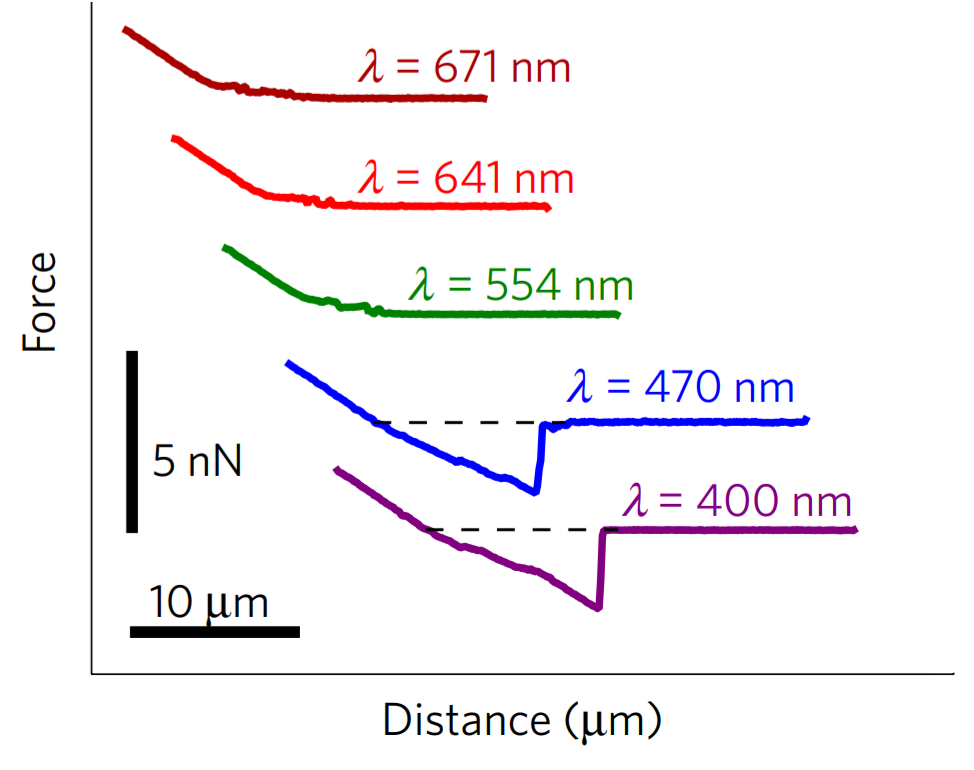
But what controls the transition? To answer this, the researchers repeated the experiment under different wavelengths of light. In Figure 5, we see that the stickiness peak is absent for red and green light but present for blue and purple light. Two potential light sensors could be responsible. One is on the cell’s eyespot and controls cell swimming to guide the cell towards the light. The other is on the flagella and controls the cell life cycle and several aspects of the cell’s mating process. But we don’t yet know which light sensor controls the stickiness, or which specific proteins make the flagella sticky.
So for the Chlamy, the decision to stay or go is made by checking if the lights are on! If they ‘go’ they can seek lighter environments, and if they ‘stay’ they can bask in the sunny spot. Watching Chlamy cells stick and un-stick as we flick a light switch is very cool, but why should we care about Chlamy? Chlamy is used in bioreactors to create biofuels and other bioproducts. Stuck Chlamy prevents light and nutrients from getting to all the cells in the reactor, so we need to understand how to control the sticking process. Plus – if we understand how a simple unicellular organism solves the problems of life, we can use this bio-inspiration for new technologies – in this case possibly new light-switchable adhesives.
[0] Should I Stay or Should I go?
[1] Antiphase Synchronization in a Flagellar-Dominance Mutant of Chlamydomonas
[2] Intraflagellar transport drives flagellar surface motility