

Original paper: Formation and size distribution of self-assembled vesicles
I’m going to start this post with an experiment. Find a piece of smooth and unwrinkled A4 or paper of a similar size, and hold it by gripping an edge between your thumb and forefinger. Due to the gravitational force, the paper is pulled down and is bent. Now crumple the same paper, then unfold and hold it by the edge again. What happened? The paper can now resist gravity! This wrinkling strategy is a simple trick to improve the mechanical response of a thin 2D sheet. Astonishingly in biology, by such simple ways, cells tune the mechanics of their thin membrane to form tiny capsules called vesicles in order to uptake nutrients, to dump waste, and to communicate. But how such a thin sheet can address all these needs? What are the mechanisms behind these tunings? Are there consequences other than mechanical improvements? In today’s paper, Changjin Huang and colleagues investigate the critical parameters governing the vesicle formation process (or vesiculation) and the size distribution of vesicles.
The Vesiculation Process
A class of molecules called amphiphiles contain two parts: a water-loving (hydrophilic) head and a water-fearing (hydrophobic) tail. When amphiphiles are dispersed in water, the hydrophobic tails are frustrated and get together (self-assemble) to stay away from water. Based on the geometry of these two parts, different structures emerge (see note [1]). One such structure is the bilayer structure (Fig. 1A).
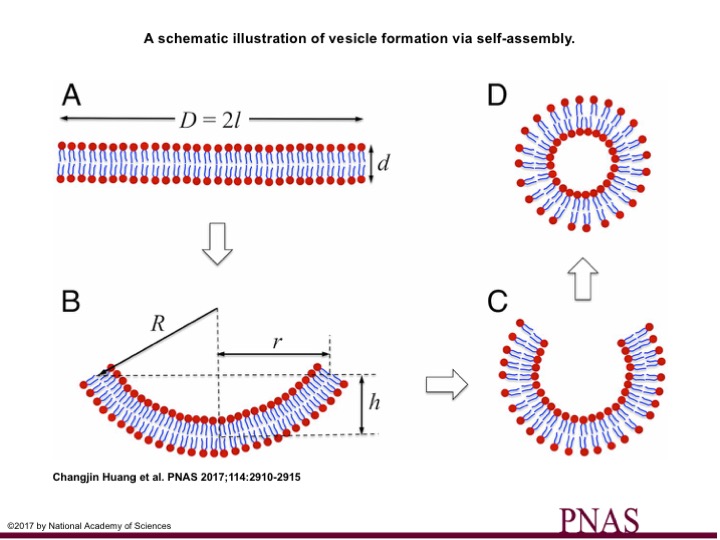
A bilayer structure composed of two layers of molecules with the hydrophobic tails turned inward (Fig 1A). This bilayer arrangement still is not the favored structure, since the water-fearing tails are exposed to water on the edges of the bilayer. An energy is imposed on the system by such exposure. This energy is called the interface energy and usually is shown by ?. This interfacial energy is the only driving force for the bilayer to bend in order to minimize the system’s energy. Thereby, the bilayer attempts to bend into spherical structures (Fig 1B & C). But bending comes at a cost! The system needs to exert force to bend the bilayer. In other words, energy is required to curve the bilayer. In this work, Huang and colleagues model this process with an energy-minimization approach to realize the critical parameters that determine the fate of this competition.
Parameters Affecting the Vesiculation
The quick paper experiment highlighted the essential role of local curvature in sheet’s rigidity, but that’s not all. The authors of this study theoretically demonstrate that besides local curvature, membrane thickness, membrane bending resistance (bending stiffness) and the membrane patch size (size of the paper sheet) all play a crucial role in the vesiculation process. When the authors considered the role of membrane thickness, they could predict morphologies other than vesicles such as disks and cups which we observe in real-life experiments.
Many models have been developed in recent decades to explain the vesiculation process, and none were able to predict the intermediate morphologies. In all of these models, the membrane is treated as a 2D sheet with no thickness such that when it is bent, only undergoes linear elastic deformation. Before we proceed, let’s briefly elaborate on ” linear elasticity”.
Imagine a spring that is being pulled by a force that you apply. The magnitude of extension is proportional to the force exerted. This example corresponds to a linear response. However, there is a threshold force after which the extension magnitude is not proportional to the force, and to predict the behavior of spring, you may need to consider non-linear terms in the model. The same consideration applies to the vesiculation process. To model the energy required to bend a membrane patch we need to consider non-linear terms since our material is a very, very thin 2D sheet undergoing an enormous deformation when bent. So, for small bendings, the small value of $latex h$ in Fig 1B, the linear term will suffice. But if membrane bends to final stages of closing itself, larger $latex h$, then we need to consider the non-linear term as well.
With this combination of linear and non-linear terms, an energy minimization model is proposed by the authors upon which a critical membrane bending length is obtained. At lengths, smaller than the critical length, the bending energy barrier increases dramatically, making it hard for the membrane to bend. At lengths larger than the critical length, bending energy barrier tends to zero and the membrane can readily bend (see note [2]). Now if we know the parameters to change this critical length, then we would be able to alter the vesicle size or to understand the mechanics of different vesicles produced by both healthy and diseased cells.
Effect of Curvature
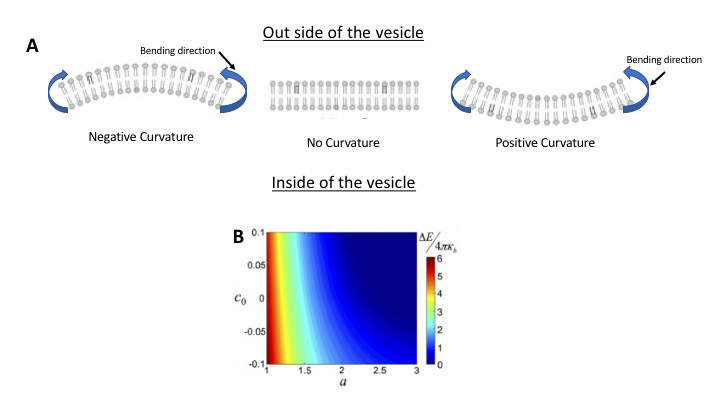
The proposed model in the original study reveals that by introducing wrinkles, we can modify the critical length, however, the model also shows that decrease or increase of the critical length by wrinkles (or membrane spontaneous curvature-see note [3]) depends on curvature direction. Under negative spontaneous curvature, the membrane is curved in the opposite direction of bending (Fig 2A). Under this condition, the model shows that the critical length is larger than when the spontaneous curvature is positive. Note how in Fig 2B, for a negative curvature the bending energy barrier diminishes only at larger critical length. So, if a given membrane bilayer has specific molecules mostly inducing negative curvature, the critical length for that membrane will be larger, meaning that the patch needs to grow more to reach the critical length after which there would be no barrier for bending. Under such condition, larger vesicles will form in contrast to the membrane with positive spontaneous curvature, which can bend itself at lower critical length, thus, forming small vesicles.
Effect of Membrane Bending Stiffness
Bending stiffness, shown by $latex k_b$ is the bending resistance of the membrane and thus it is a membrane property. Sometimes cells recruit molecules such as cholesterol to their membrane to increase the membrane bending stiffness. On the other hand, viruses are known to decrease the membrane stiffness so that they can readily bend the host’s cell membrane. In regard to vesicle size distribution analysis, the proposed model showed that the critical length is proportional to bending stiffness. In other words, for the stiffer membrane, the critical length is larger and these membranes tend to form larger vesicles.
Effect of Membrane Thickness
So far, for our analysis of the membrane (or sheet for our analogy) thickness was fixed. To consider the membrane thickness, the authors adopt a simple approximation. They first argue that membrane stiffness varies as a function of membrane thickness squared ($latex k_b \propto d^2$). Then, assuming that membrane is free to bend (its size is larger than the critical length), they obtain the minimum diameter of the vesicle formed from this membrane size as $latex D_{min}=(critical\ length) + (membrane\ thickness)$. But $latex critical\ length \propto k_b$. Therefore, from their argument we can write:
$latex D_{min}=d^{2} + d$
With this approach, membrane thickness is considered as a non-linear concept. The proposed model reveals that for thicker membrane the critical length is larger, and thus these membranes will more likely form larger vesicles. In contrast, for the thinner membranes, the critical length is shorter and these membranes are prone to form small-sized vesicles.
Conclusion
The vesiculation model developed by Huang and his colleagues has contributed to our understanding of how vesicles form. Understanding the parameters that govern vesicle formation is critical for the design of vesicles for applications such as drug delivery, where nanoscale vesicles are needed to move drugs into a cell. In addition, the identified vesiculation parameters could be used as diagnostic measures, as it has been shown that the vesicles produced by cancer cells or by cells infected with viruses have mechanical properties different from healthy cells.
[1] Known as Israelachvili’s packing parameter, the volume of the hydrophobic part divided by the product of effective hydrophilic area and the length of the hydrophobic part,$latex p=\dfrac{v}{l*a}$, defines the favored morphology. when p < $latex \frac{1}{3}$ spherical micelles, $latex \frac{1}{3}$ < p < $latex \frac{1}{2}$ cylindrical micelles, p > $latex \frac{1}{2}$ bilayer structures are expected to form.
[2] Cut an unwrinkled A4 paper in half and see the bending response. If you continue cutting you will notice that after a certain length the paper doesn’t bend. That length is the critical length.
[3] Spontaneous curvature is the natural curvature of the membrane because of asymmetries between two monolayers of the bilayer. These asymmetries can be due to the presence of proteins or geometrical difference of different amphiphilic molecules making the membrane.