Original paper: Emergent Sensing of Complex Environments by Mobile Animal Groups
Imagine you forget to bring money for lunch, and you overhear a teacher mention that there is free pizza somewhere on the third floor of your school. If you’re alone, you might walk around the third floor, trying to detect signs of pizza – does a room smell delicious? Do you see a suspicious stack of pizza boxes by the door to the gym? Just by using your senses, you can find the pizza. However, it is likely that there are other students on the third floor who also want free food. Maybe if you follow a crowd of students all walking in the same direction and talking about whether they want a Hawaiian or pepperoni slice, they might lead you directly to the pizza!
Which of these methods will be more effective? Following environmental signals, such as the smell of cheese, or social signals, such as the people all heading in the direction of potential pizza? In “Emergent Sensing of Complex Environments by Mobile Animal Groups,” Andrew Berdahl and colleagues seek to find out how searching in groups enhances the sensing ability of animals.
The researchers used a fish called the golden shiner to study this kind of mob mentality. These fish live in large schools in shallow water and prefer darker habitats. Fish school together for many reasons. For example, it helps them avoid predators and gain advantages in hunting. In this experiment, the researchers asked whether schooling helps the fish find their preferred darker spots in the water. A school of golden shiners searching for dark spots in water is a convenient model system, but the researchers stressed that the results from this study can be applied to any group of organisms looking for any environmental cue.
Berdahl and his colleagues set up a large, shallow tank for fish to swim in. The tank was in a dark room, and a projector was used to impose light patterns on it. The patterns consisted of a bright tank (similar to an overcast day) with dark patches (similar to twilight). The dark patches moved around randomly at a constant speed, with the fish expected to follow the patches.
Fish were tested as individuals and as groups from 2 to 256 fish. To track the fish in both light and dark regions, the researchers used infrared (IR) light that the fish can’t see and took videos of the fish with an IR camera. The fish could then be tracked using image analysis. You can see the visible light and IR images of the fish in a dark spot in Figure 1.

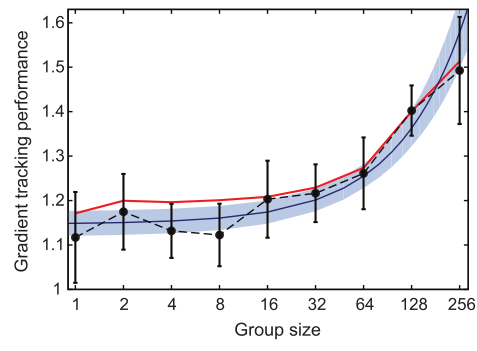
How well the fish stayed in the dark was measured with a performance metric, $latex \Psi$ (psi). This number measures how good the fish are at staying in the dark. Specifically, it measures the average inverse of brightness at all the fish positions averaged over time [1]. If $latex \Psi = 1$, the fish did not try to stay in the dark at all; the performance was better as $latex \Psi$ increased. The data in Figure 2 shows that $latex \Psi$ increases with group size – bigger groups make the fish better at tracking dark patches.
The researchers wanted to find out whether the fish were responding to the changes in the environment or the behavior of their neighbors. They calculated a social vector and an environmental vector for each fish. The social vector measures what direction a fish’s closest neighbors are. If all of the fish’s neighbors are to its left, there will be a strong social vector to the left; if the neighbors are all spread out around it the social vector will be very small [2]. The environmental vector points in the direction of the darkest position near the fish[3]. The researchers calculated how correlated each vector was with the acceleration of the fish. When the magnitude of the social vector was very high – a fish’s neighbors were all located in the same direction from it – the fish listened to the social vector and swam to where their neighbors were. They did not respond as strongly to what direction the nearest dark patch was in. In other words, fish respond much more to nearby clusters of neighboring fish than to their environment, similarly to how you might pay attention to your friends in your hunt for pizza rather than smelling around.
Although fish did not respond to the changes in the lighting of the tank directly, as measured by the environmental vector, they did respond to the environment: fish swam faster in lighter regions, and slower in darker ones. They responded to the scalar brightness at their position in the tank, rather than how much the brightness was changing. If a mountain climber behaved like these fish, he would climb more slowly at higher altitudes (responding to the elevation) but not change his speed based on the steepness of the slope (not responding to the gradient in elevation).
These two main behaviors of the fish, swimming towards their neighbors and changing their speed based on the lighting, made group tracking of dark patches very effective. The researchers highlight two examples of how this could work. The first example is that of fish traveling next to a dark patch. Some of the fish are located on the brighter side of the dark patch and swim faster. Other fish are in the darker region and swim slower. This causes the whole group to turn towards the slower fish and therefore into the dark patch. The second example is of a group of fish traveling into a dark patch. As fish enter the dark patch, they slow down. The rest of the group follows them and slows down as well. This increases the number of fish in darker regions.
The researchers created a computer model that simulated behaviors of fish with these two rules: following their neighbors and changing the speed according to the brightness of the light at their positions. Although they did not build in any explicit response to the changing light gradient, the groups of simulated fish responded the same as the real fish in the experiment, as seen in Figure 2. Berdahl and his colleagues conclude that the response of the fish to the environment arose simply from those two rules, and the ability of groups to track dark patches increased with larger numbers of fish.
The researchers emphasize many times that the results of this study are applicable to any group sensing any field, not just fish in a light field. The results could apply to bacteria seeking food, or robots seeking a resource to collect. If, for some reason, a group of animals is broken up – for example, there are fewer fish in a school due to overfishing – the remainder of the fish in the school might not be as well equipped to seek out darker patches to hide from predators. This study highlights the importance of paying attention to your neighbors and the advantages all living organisms gain from working in groups – like helping a hungry student find some pizza!
[1] The performance metric is defined as:
$latex \Psi=\frac{\langle \langle 1-L \rangle_{fish}\rangle}{\psi_{null}}$
where L represents the light level and $latex \psi_{null}$ normalizes $latex \Psi$ so that $latex \Psi = 1$ implies that fish do not track the light at all. The inner angle brackets represent the average of the darkness, $latex 1- L$, taken over all fish, and the outer angle brackets represent the average taken over all time
[2] The social vector is defined for each fish as:
$latex S_i=\sum \frac{c_j-c_i}{|c_j-c_i|}$
where ci represents the position of the ith fish, so cj–ci is the difference between the positions of the ith fish and its neighbor, the jth fish. It is normalized by the magnitude of that distance. The sum is taken over all the fish within seven body lengths of the ith fish, for each fish
[3]The environmental vector is the negative of the gradient of the light field for each fish, i:
$latex G_i = -\nabla L\mid_i$