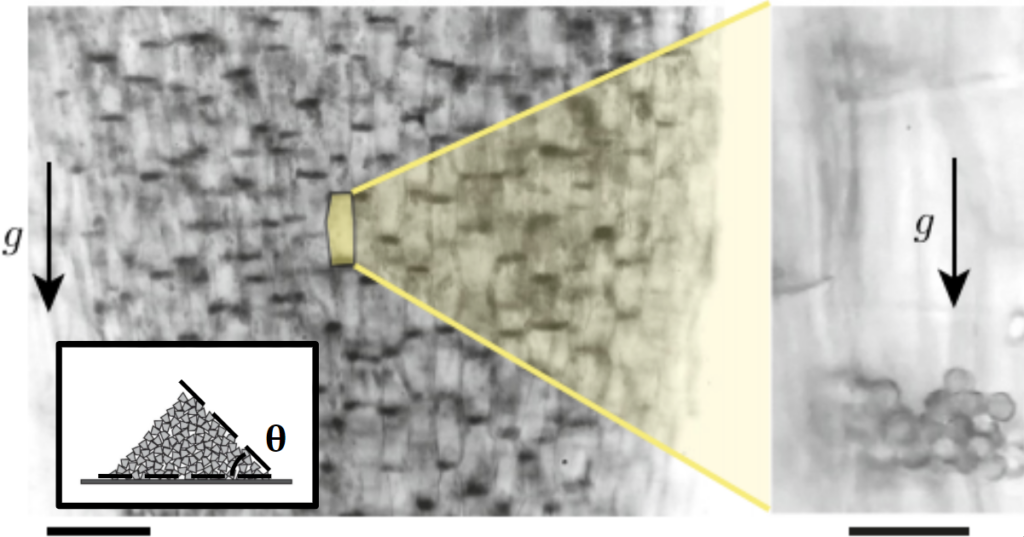
Plants need to know the direction of gravitational pull in order to grow their roots downward and their stems upward. This information is crucial whether the plant grows in your garden, on a cliffside, or even on the International Space Station [1]. While it’s been said that it took a falling apple for Newton to figure out how gravity works, our photosynthetic friends use a more intricate microscale sensor to detect gravity. This sensor consists of starchy granules called statoliths which can be found on the bottom of specialized cells called statocytes. An accumulated pile of around 20 statoliths at the bottom of a statocyte cell is shown in Figure 1. If the cells are tilted like in Figure 2, the pull of gravity initiates a statolith avalanche that indicates the direction of gravity. The position of statoliths is part of a complicated signaling network that tells the plants how to correct its growth towards or against the direction of gravitational pull.
Figure 1. Microscope images of statolith piles in gravity-sensing statocyte cells of wheat coleoptiles, which are the sheaths covering an emerging shoot of wheat. Arrows indicate the direction of gravity. (Left) Statolith piles are visible as dark areas on the bottom of the cells. The scale bar represents 100 µm. (Right) Close-up view of the statolith piles. The scale bar represents 20 µm. Images courtesy of the original article. Inset: Standard granular pile just below the avalanche angle. Schematic courtesy of Andreotti et. al., Granular Media Between Fluid and Solid. Figure 2. Movie made from microscopic images of statolith avalanches in wheat coleoptiles after the cells are tilted 70 degrees. Statolith piles are dark spots. Movie runs at 40x speed for a total duration of 10 real minutes. Courtesy of the original article.
However, Bérut et. al. realized that this description of statolith piles didn’t totally agree with our knowledge of granular materials. There are two major issues. First, granular piles are known to initiate avalanches only when the slope of the pile reaches a critical angle, usually between 5° and 30° depending on the characteristics of the grains. In the case of the statoliths, the critical angle was found to be around 10°. When the slope is lower than the critical angle, the pile should be completely immobile due to frictional forces between the grains. However, plants are able to detect even the slightest changes in gravity — involving angles much smaller than 10° — indicating that avalanches are not the whole story. Secondly, upon tilting as in Figure 2, the grains seem to avalanche until they establish a flat surface layer. This is in direct contrast to classical granular materials. If statoliths behaved classically, we would expect them to avalanche until the critical angle of 10° is reached, rather than their actual final angle of around 0°. How can we explain the shocking sensitivity of these granular piles?
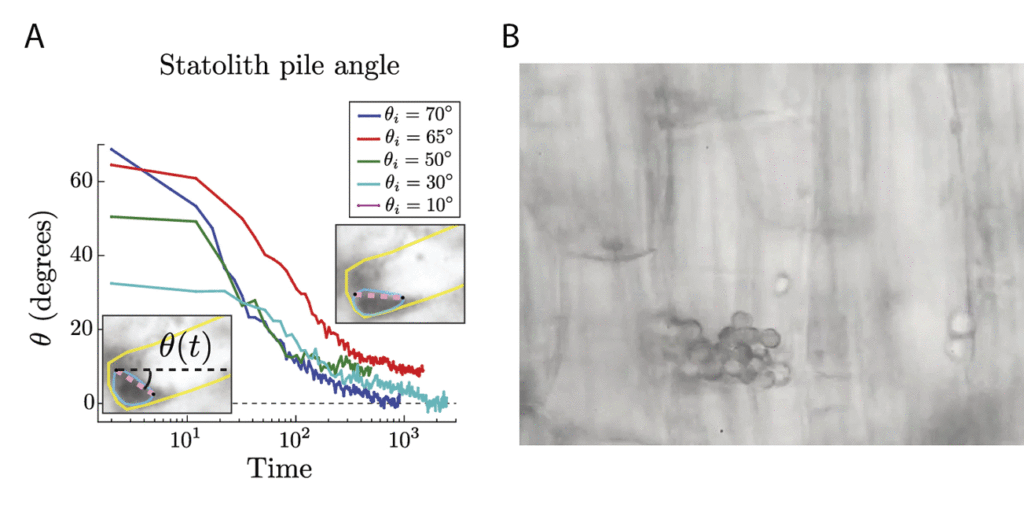
Figure 3. (A) Two observed dynamical regimes in the averaged angular decay of statolith pile slopes over time. Each curve corresponds to different initial inclination angles. (Inset) Initial and final configurations of a statolith pile after being tilted by 70°. (B) Close-up video of a statolith avalanche (when tilted 15°) highlighting the random motion of individual statoliths. Movie is played at 80x speed, for a real duration of 14 minutes. Images and video courtesy of the original article.
By studying the flow response of the statoliths to gentle inclinations, Bérut et. al. found that the statoliths in fact flow, liquid-like, from a pile into a puddle with or without prior avalanches! Figure 3A shows a statolith pile angle slowly creeping from 10° (or less) to 0° in 10-20 minutes. This creeping response occurs at any slope. Under the microscope, the statoliths are seen to vibrate, with each individual statolith undergoing random motion shown in Figure 3B. The statoliths are agitated, the origin of which is likely biological processes within the cell rather than random thermal motion, as thermal energy is too small to drive the observed grain activity. While we know that classical granular piles do not flow below the critical avalanche angle, this is not the case for active granular materials. Agitation allows the grains to free themselves from the pile, turning an otherwise static mountain into a fluid-like substance. Long before we had an understanding of the physical world, nature was already building and refining amazing biological machines. We have only recently begun to understand the properties of agitated granular materials, meanwhile plants have been using active grains to detect gravity all this time. Given plants’ long-time expertise with gravity, perhaps the apple that fell on Newton’s head was nature’s way of telling humans to hurry up and figure it out.
Evolution usually solves challenges differently than human engineers—something easy for biology is often difficult for us, and vice versa. Learning from biology can help us solve difficult challenges more easily. One example of this is making complex optical lenses.
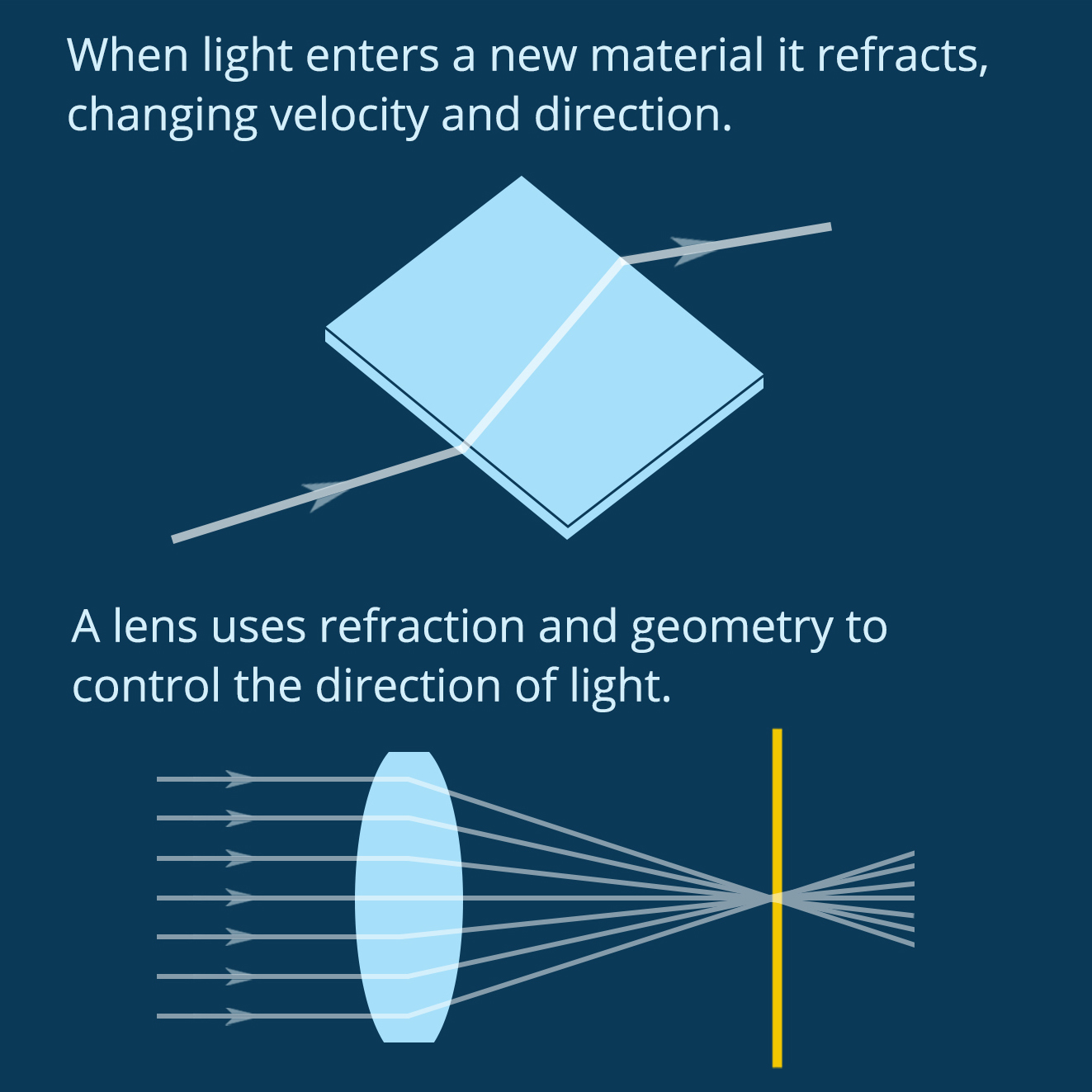
Figure 1. Lenses use refraction and geometry to direct light. (Figure by the author)
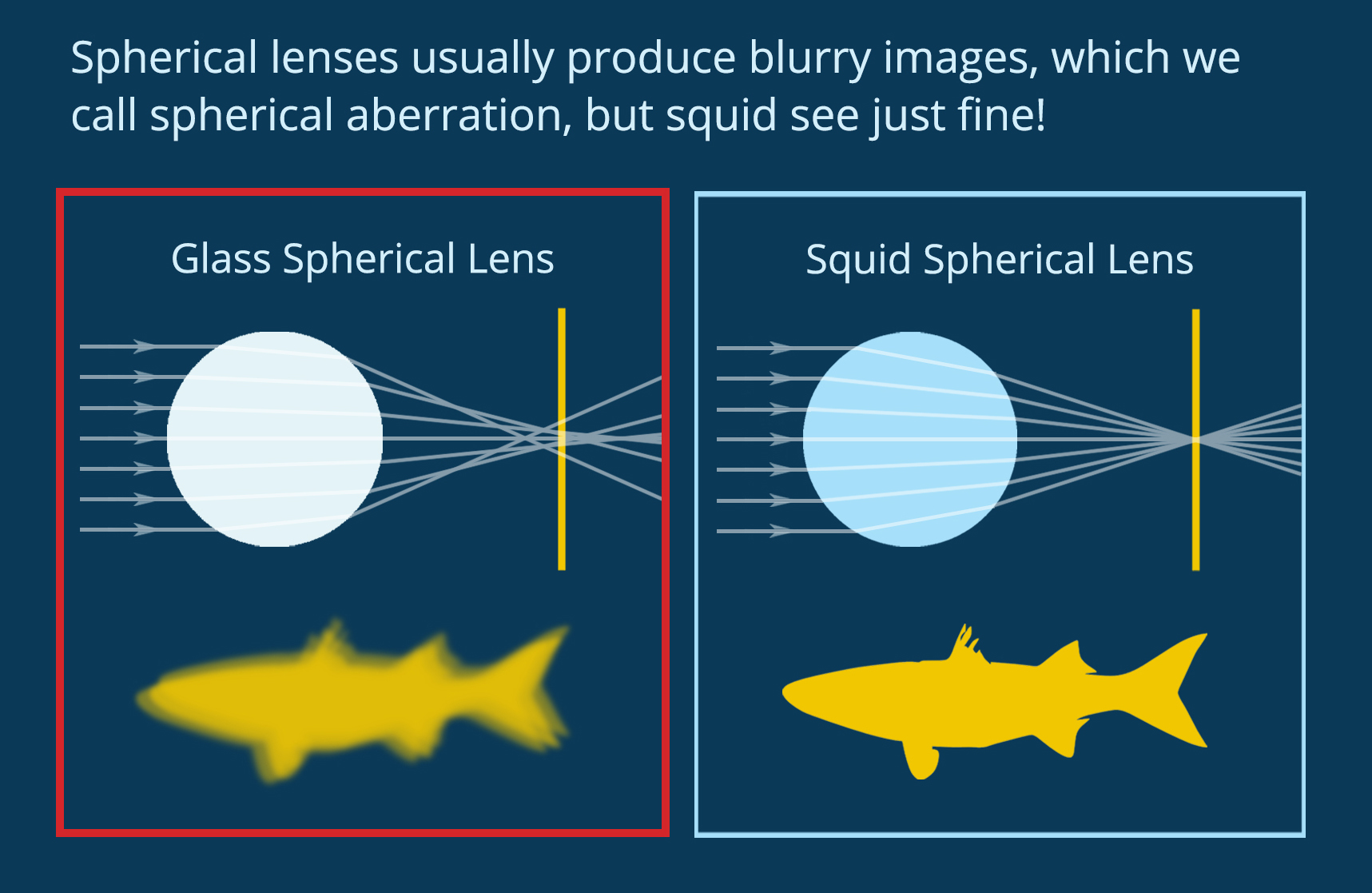
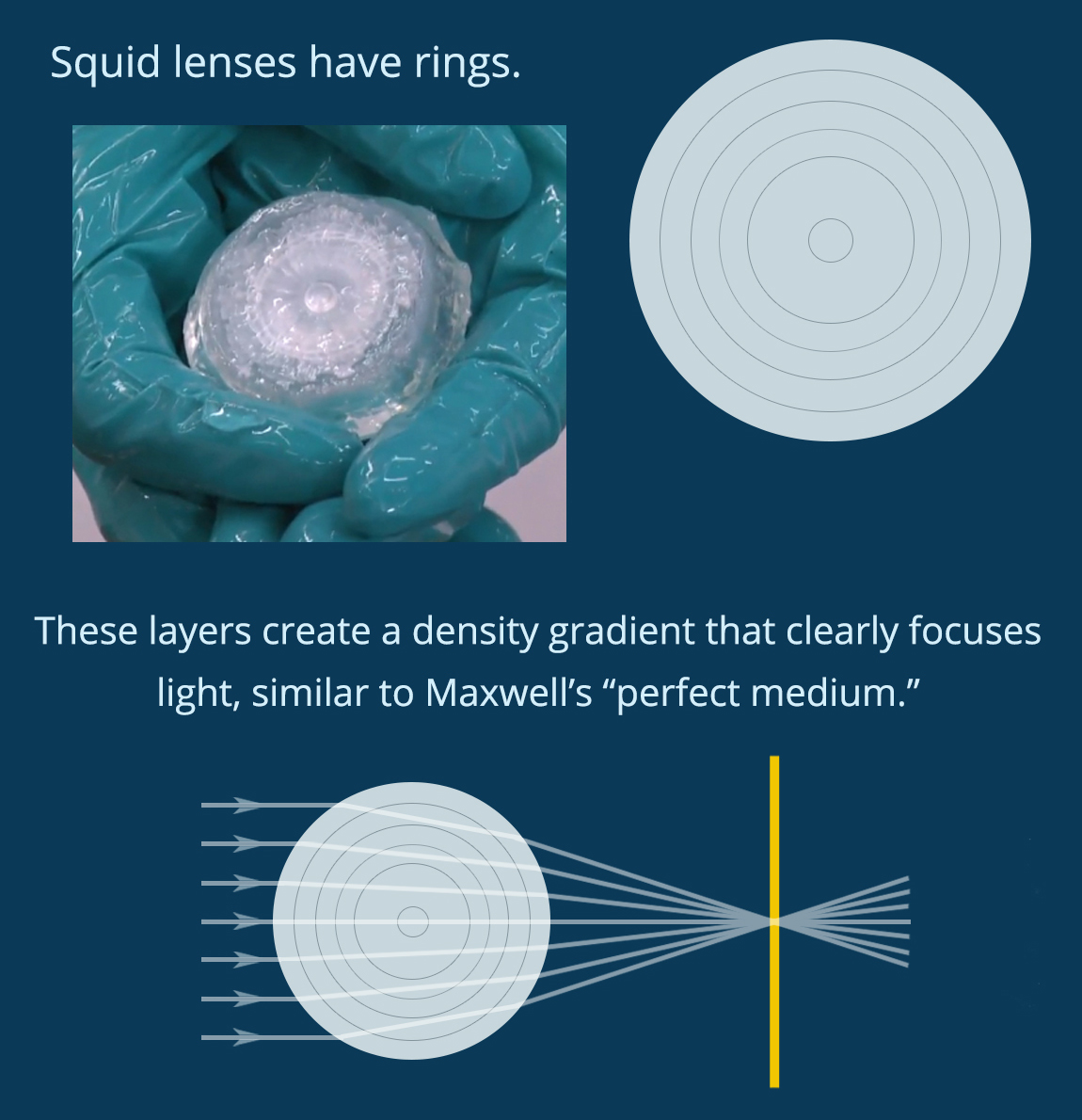
When light enters a new material it refracts, changing velocity and direction. A lens uses geometry and material properties to direct light on a specific path. You can see in Figure 1 how the shape and refractive index of a biconvex lens combine to direct light at a single spot. In biology, complex eyes like those found in most vertebrates and in squid have a lens that directs light onto the retina at the back of the eye, forming an image to be processed by the brain. Squid use spherical lenses to do this, but spherical lenses have a problem. As you can see in Figure 2, if you make a spherical lens out of one material (like glass), the light rays overlap after exiting the lens and the resulting image is blurry. This is called “spherical aberration.” Human engineers use spherical lenses a lot, and we correct for spherical aberration by combining multiple lenses. Squid, on the other hand, have evolved a lens that self-corrects for this distortion.
Figure 2. Spherical lenses usually produce blurry images, but not in squid. (Figure by the author)
We know, in theory, how a squid might do this. In 1854, the famous physicist James Clerk Maxwell mathematically designed a spherical lens with “perfect” focus. He showed that if the density of the lens changes along the radius, forming a density gradient that he called a “perfect medium,” then the lens will produce a clear image. Today engineers can make gradient index lenses like this, but the process is difficult and energy intensive. Squid evolved to grow them easily. Could understanding how squid make these lenses help human engineers learn to do the same thing? This question inspired Dr. Jing Cai and Prof. Alison Sweeney to study the structure of the squid lens.
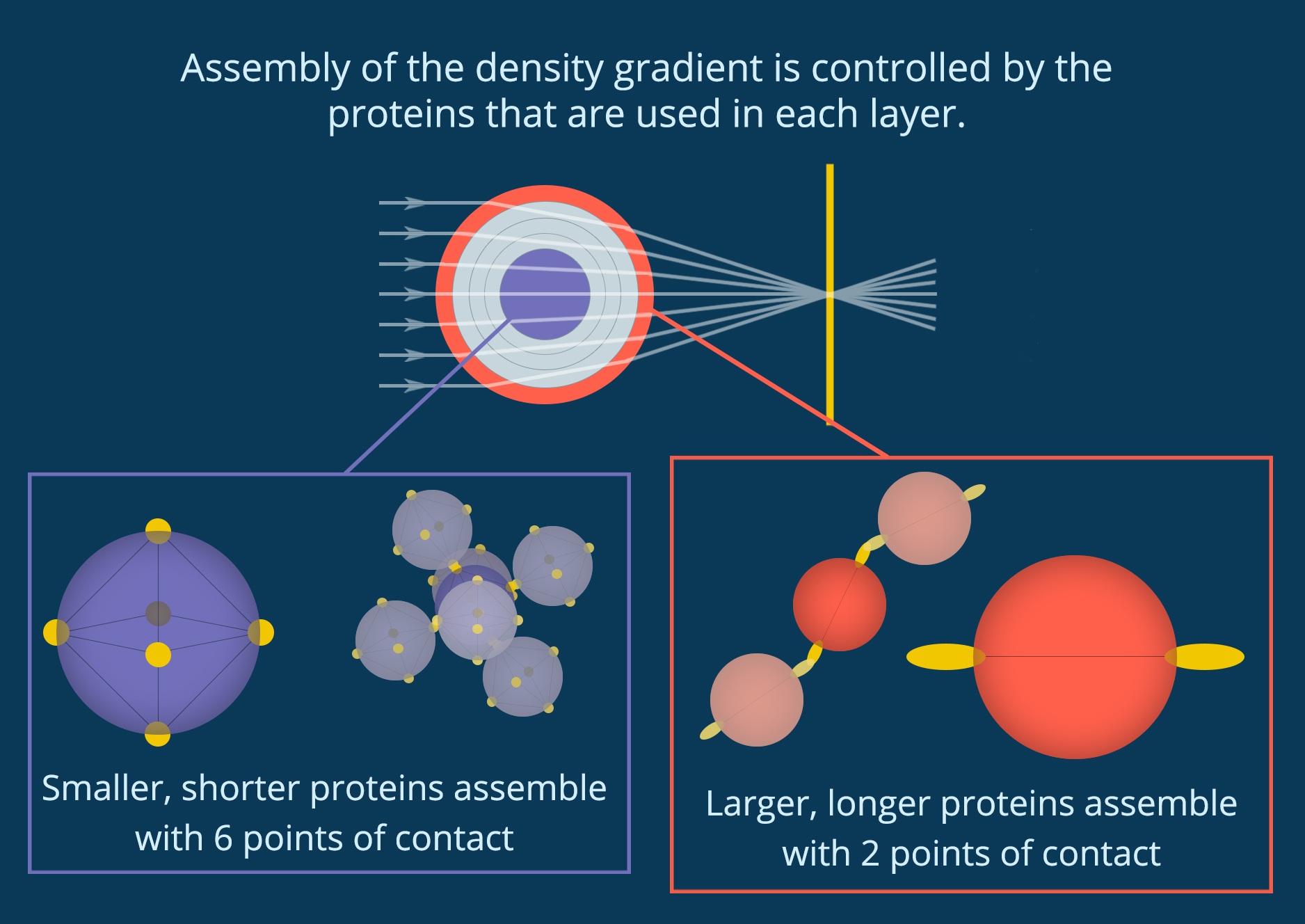
You can see in Figure 3 that when you crack open the lens in a squid eye you find rings like the inside of a tree trunk. If each of these rings has a slightly different density, they could combine to create a perfect medium. The lens of an eye is made out of proteins called crystallins, which fold into individual particles before linking together into a single material. Cai and her collaborators discovered that the lenses of the Longfin inshore squid (Doryteuthis pealeii) use 53 different crystallin proteins of different sizes. They also found that the different proteins are used in different parts of the lens, and each layer of the lens has a slightly different structure. As you can see in Figure 4, small proteins at the center of the lens are densely packed together so that each protein is connected to six other proteins. However, the larger proteins at the edge of the lens have more space between them, and each protein only touches two others.
Figure 4. Protein particles assemble differently in different parts of the lens, creating a “perfect medium.” (Figure by the author)
This makes sense when you think about the cells that make these proteins. Cells rely on diffusion to bring building blocks to the right place for protein assembly and to send each assembled protein out to where it’s needed. When finished proteins link together to grow the lens, they disrupt this diffusion and stop protein production. By growing from dense to less dense and using so many different proteins (53 in the Longfin inshore squid), the cells are able to start and stop the growth of different layers while maintaining a single particle network. No part of the lens separates out or turns opaque, but there are still large enough regions with different densities to diffract light into alignment.
Cai and her collaborators showed that squid lenses definitely use a density gradient similar to Maxwell’s perfect medium to correct for spherical aberration. It’s likely that this density gradient not only creates a perfect medium, but also helps control lens assembly. Now that we know how squid build a perfect spherical lens, it is easier to envision how human engineers could grow our own complex optical materials.
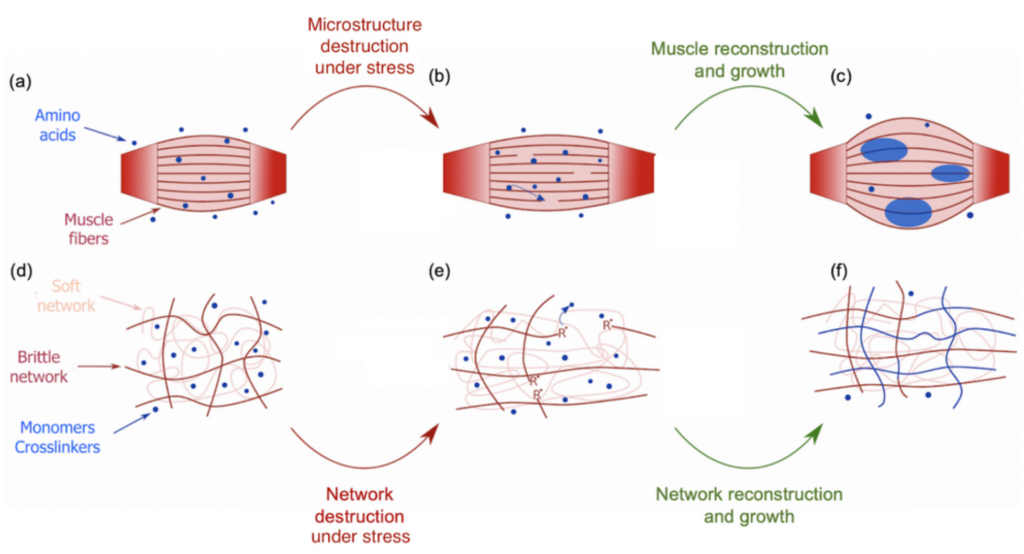
During the COVID-19 lockdown, many of us had the opportunity to workout, for instance by lifting weights to build stronger biceps. During a workout, our muscles undergo some damage at the micrometer scale that triggers an immune system response (Figure 1a). Adequate amino acids, the main constituents of proteins that give muscles their structure, are then carried close to the torn tissues to repair the damage and grow new muscle thanks to the binding between the amino acids and the tissues [1]. This process is a sort of mechanical solicitation where your body creates stronger biological materials with more mass (Figure 1b-c). This seems, however, counter-intuitive when compared to the response of synthetic soft materials under the same mechanical solicitation. Indeed, mechanical stress usually weakens or even damages synthetic materials. For instance, pulling too hard on an elastic band will result in its irreversible failure. Can we then use muscle growth as an inspiration to design materials that would get stronger and bigger under mechanical stress? By mimicking this process, a group of scientists from Hokkaido University developed such a material. This new material belongs to the family of polymer hydrogels. Hydrogels consist of a stretchable 3D network of polymer chains in water connected to each other by molecules called crosslinkers. These crosslinkers control the stiffness of the network. Indeed, a highly connected network will require more stress to be deformed than a loosely connected one.
The difference is that this new material is a double network hydrogel — a hydrogel with two interpenetrated networks — one of them is brittle and rigid due to a high content of crosslinkers, shown as red chains in Figure 1d, while the other one is soft and stretchable due to low content of crosslinkers, shown as pink chains in Figure 1d. This double network is immersed in a solution containing 80-90% water and two types of molecules that are the building blocks of the hydrogel: monomers and crosslinkers. When a tensile stress is applied to the material, the brittle network is the one that “feels” the pressure, leading to breakage of its strands while leaving the soft network undamaged (Figure 1e). During the failure of those polymer chains, the bonds between the carbon atoms break down due to the stress which generates highly reactive chemical species known as radicals [2]. These radicals initiate the formation of new polymer chains by reacting with the building blocks in the solution [3]. As these new chains are also connected by crosslinkers during the reaction, the damaged network is then not only restored but extended and strengthened [4].
Figure 1. Illustration of the bio-inspired study on strengthening double network hydrogels under mechanical stress. Top: a muscle initially at rest (a), after being damaged under effort (b) and reconstructed and grown by the action of neighboring amino acids (c). Bottom: a double network hydrogel at rest (d), after being damaged by tensile stress (e) and reconstructed and grown by the action of neighboring building monomers and crosslinkers (f)
The most remarkable result which validates this process is shown in the following movie :
Movie 1. Cyclic test of tension and recovery of a double network hydrogel caught between two clamps. The setup is immersed in a solution of monomers and crosslinkers suitable for reconstruction and growth of the material after mechanical solicitation.
A double network hydrogel is stretched at constant stress three times in a row. The hydrogel is immersed in a solution of monomers and crosslinkers that will be used up for the brittle network reconstruction. After each cycle of stretching, a one-hour reconstruction and growth period allows the material to strengthen. Thanks to that stretching and recovering protocol, the material is able to lift a 200g weight higher and higher as it gets stronger.
The successful emulation of muscle self-strengthening applied to synthetic materials design paves the way for tremendous new bio-inspired materials. They are of especially great interest in soft robotics, where soft materials are exposed to harsh conditions that may lead to damage, like cuts. Rather than undergoing simple regeneration, the material would respond to damage by strengthening to better resist its environment.
[3] This polymerization reaction is called radical polymerization. The radicals, R, in Figure 1e will react with the monomers which contain chemical groups sensitive to radicals (double carbon bonds for instance). The result of this first reaction is another radical which will react with another monomer, thus, by repeating the process, connecting the monomers into a polymer chain.
[4] The generation of enough radicals and their contact with monomers and crosslinkers are assured by the second soft network which maintains the cohesion of the whole material while bonds are broken. This is comparable to a muscle where the breakage of its microstructure does not provoke the collapse of the entire muscle.
Quintessential soft matter problems, such as the behavior of droplets in ink-jet printing, involve complex interactions between forces and materials. In today’s article, Prof. Wilson Poon points out that coronaviruses are also quintessential soft matter objects, and highlights a range of areas where soft matter science may help better understand, and combat viral pandemics.
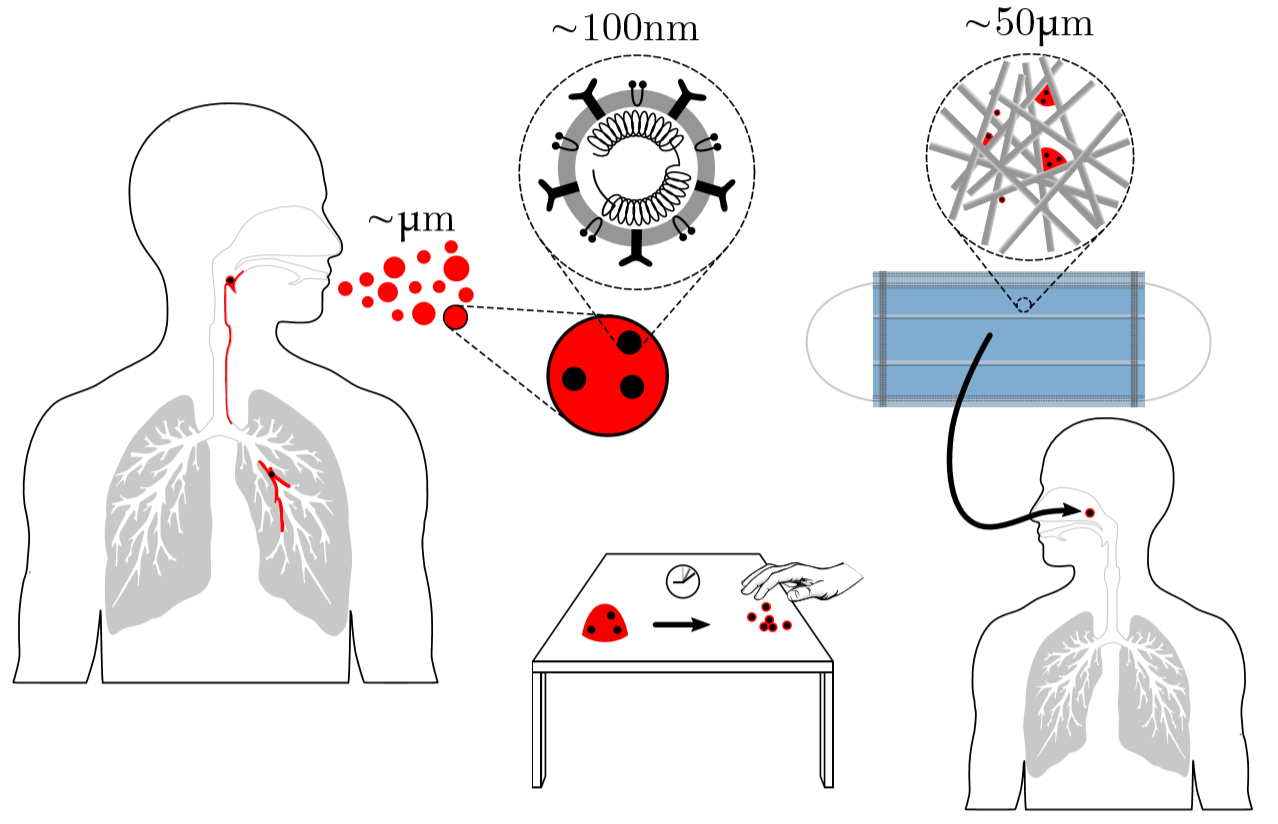
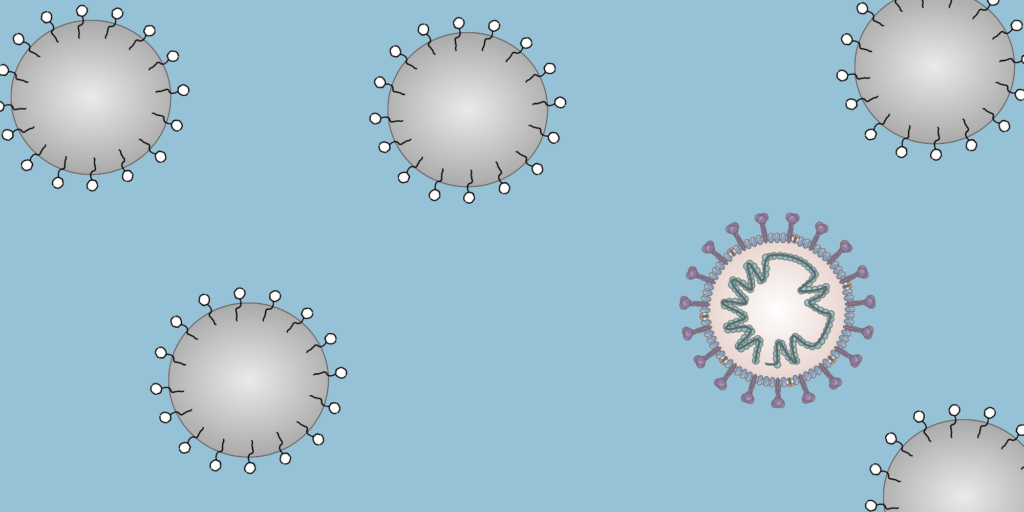
To a physicist, a virus is merely an inert particle that drifts around in water (it is a colloidal particle, in the jargon of soft matter physics, see Figure 1), where a single coronavirus particle is roughly spherical and around one tenth of one thousandth of one millimeter in width ($latex 0.1 \mu m$). These inert particles, however, are also covered in tiny specially shaped keys (proteins), and when these keys meet the correctly shaped lock (a receptor at the surface of a cell membrane), they activate. The previously inert particle now instructs the invaded cell’s machinery to make more viruses (imagine a Ford car whose only job is to drive into Rolls Royce factories and rewire the machines to make more Fords). Sickness is then caused by the cells being too busy making viruses to do their jobs, eventual cell death, vast numbers of obstructive virus particles, and the body’s own army of immune response units.
Although less interesting biologically, while in the inert stage the virus has to “survive” a variety of physical conditions, and ideally this is where we want to destroy it, before it has a chance to invade a cell — before the Fords can find the Rolls Royce factories. In this article, we will focus on a number of open questions about coronaviruses prior to invading a host that soft matter could help answer.
Figure 1. Diagram of a colloidal solution, particles of diameter 0.1 micron. Most are standard surfactant coated solid particles used widely in industry and academia, whereas one is a coronavirus.
Airborne
It is now established that the “dominant route” by which coronaviruses spread is in airborne droplets, through sneezing, coughing and even speaking. In soft matter physics, airborne droplets have been widely studied, both in industry (for example in ink-jet printing) and in academia (for example in finding the scaling laws that govern droplet formation). We may be able to use the lessons we have learned to ask:
How are these droplets created?
Does the behaviour of these droplets impact virus survival and transmission?
Do all droplets contain the same number of active virus particles?
Can we modify the air to inhibit viral transmission via the behaviour of these droplets?
When sneezes are observed using high-speed photography, as in this video, an image of a turbulent cloud of droplets is revealed. These droplets break apart, collide, stretch and break apart again in a chaotic cascade that could exert considerable destructive shear forces on the viruses, but it is currently unknown whether the turbulence decreases the active virus concentration or if the virus particles influence the droplet break-up events. Solving this unknown may change the way we understand the role of air flows from ventilation systems in inhibiting active viruses.
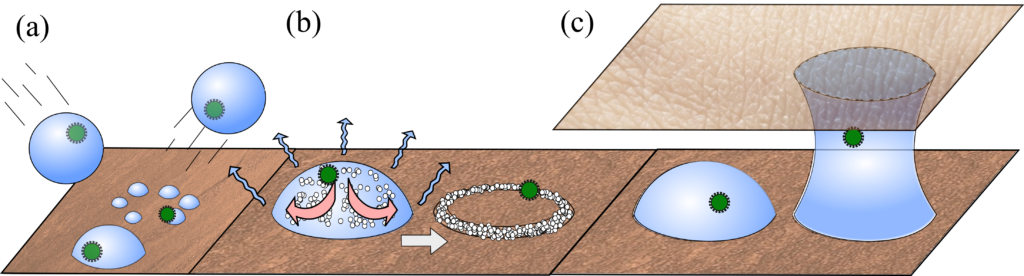
Despite the lack of information on virus viability in the turbulent sneeze cloud, Poon and colleagues point out that studies suggest that the viral load of virus-carrying droplets is small – around 1 viable virus particle per droplet. However, sneezes typically generate up to 40,000 droplets, whereas a single cough (or talking for 5 minutes) will generate up to 3000. One also needs to know how many viruses a person must absorb before their dose becomes infectious – the minimum infective dose (MID). For the flu, the MID is reported to be around a few thousand, but the equivalent figure is not known for coronaviruses. All of these numbers play a role in infection but collectively point to the importance of limiting the amount of direct droplet transmission, and the importance of ventilation.
Finally, an airborne droplet containing a virus evaporates, at a rate depending on the local humidity, and will eventually completely dry. Interestingly, studies suggest that bacteriophages (viruses that invade bacteria) are more likely to survive in very dry or very humid air, but not in between. The reason for this seems to lie in how dissolved salt destructively interacts with the virus at steady (not slow or fast) evaporation rates. Other saliva components (like the gelation of the mucin proteins that make mucus slimy) also impact evaporation, further complicating the problem of viral transport and survival, but these clues indicate that controlling air humidity could be a viable option for hampering the virus.
On Surfaces
In the previous section, we’ve seen that how droplets behave in the air seems to be important, and this has been well-studied in a variety of commercial and academic contexts. As in ink-jet printing, we should also consider how droplets impact surfaces, as contact from surface to skin to mucus-membrane is another route through which viruses spread.
Here we will summarize the five main questions highlighted by Poon and colleagues in how droplets interact with surfaces, which could be important for understanding viral survival and transmission:
Splashdown. While ink-jet printed droplets need to avoid bouncing off surfaces, in a pandemic we ideally want the opposite. Polymers are known to give an anti-bounce property to droplets, but do mucins in saliva also have this effect? (see Figure 2 a)
Coffee rings. Much research has been done on the coffee-ring effect (When Espresso Evaporates), where suspended material is dragged to a droplet’s edges as it dries. Do viruses cluster at the edges in this way, and do these dragging forces impact their viability? This is further complicated by the salts and mucins present in these droplets, which will also accumulate at the edges (see Figure 2 b).
Material. Surfaces can have complex structures (smooth, ridged, or fibrous) and variable chemistry (metal, glass, or oily skin cells). Interestingly, virus viability tends to follow an exponential decay once on a surface, but the decay rate is higher on some surfaces (e.g. copper) than others (e.g. plastics). A question remains: is this due to a chemical catalytic property, or are some surfaces retaining moisture better than others?
Capillary Bridges. Watery bridges can form when two wet surfaces come into close contact, but do microscopic bridges form when your skin comes into contact with surfaces, and do they enhance viral transport? Interestingly, bacteriophages are known to better transfer from surface to surface in humid environments, which suggests that such liquid bridges may be important (see Figure 2 c).
Fluidic Forces. It is well documented that bubbling gas through a solution or filtering it through a bed of glass beads deactivates viruses, but why? Poon argues that there could be significant capillary forces (which are known to deform latex particles in drying paint) pulling on virus particles if they get stuck at the interface between air and water, which will likely happen often in a vigorously bubbled or filtered solution. The behaviour of colloidal particles at interfaces is a rich area of ongoing study, so answers here are anticipated soon.
Figure 2. Schematic of processes to consider with viral loaded droplets on surfaces. (a). Droplets impacting surfaces. Bottom: droplet impact is absorbed, and it comes to rest in a single spherical cap. Middle: Droplet splashes into multiple droplets. Top: Droplet bounces away. (b) Droplet evaporation. Particles, salts, mucins and viruses accumulate at droplet edges via the coffee-ring effect. (c) Capillary bridges. Schematic shows a pathway viruses may take via capillary bridges between the resting surface and skin.
Final Note
The possibility of a global viral pandemic had been predicted by scientists for decades, and yet was not prevented, and the current one is seemingly unlikely to fully disappear in the near future. While the world’s hopes currently rest with developing and deploying effective vaccines, we should in parallel use every single weapon at our disposal to understand and treat the causes and symptoms of viral pandemics. The lessons we learn may also allow us to improve on some of the problems that the current measures are creating, such as the unfortunate and unsustainable use of single-use plastics in disposable protective gear that contribute to the “plastic pandemic”. Soft matter science may not be central to solving the problem of COVID-19, but is important, and could have a significant role to play in preventing the next global pandemic.
Featured image for the article is an edited version of a figure from the original article.
If you speak to a soft matter physicist these days, within a few minutes the term “active matter” is bound to come up. A material is considered “active” when it burns energy to produce work, just like all sorts of molecular motors, proteins, and enzymes do inside your body. In this study, the scientists are focusing specifically on active polymers. These are long molecules which can burn energy to do physical work. Much of biological active matter is in the form of polymers (DNA or actin-myosin systems for example), and understanding them better would give direct insight into biophysics of all kinds. But polymers are microscopic objects with complex interactions, making them difficult to manipulate directly. To make matters worse, physicists have yet to fundamentally understand the behaviors of active materials, since they do not fit into our existing theories of so-called “passive” systems. In this study, Deblais and colleagues decided to entirely circumvent this problem by working with a much larger and easier-to-study system that behaves similarly to a polymer solution: a mixture of squirming worms in water.
The researchers focused on the viscous properties of this living material, which behaves somewhat like a fluid. Viscosity is a measure of a fluid’s resistance to gradients in the flow. Polymer fluids are highly viscous because the long molecules in a polymeric liquid get tangled up in one another. Physical descriptions of most fluids assume that viscosity is a constant (so called Newtonian fluids), but many materials exhibit what is called shear thinning. This is when a fluid flows more easily as one applies an increasing shear force, that is, a force pulling the system apart. We encounter shear thinning at the dinner table all the time when struggling to pour ketchup, another polymeric fluid, out of a bottle. If the bottle is shaken fast enough, increasing the shear force applied, the ketchup flows smoothly like a liquid. In polymer systems (like xanthan gum in the ketchup) shear thinning happens when polymers are pulled apart fast enough that they tend to align together, which loosens the entanglements that held the system together before.
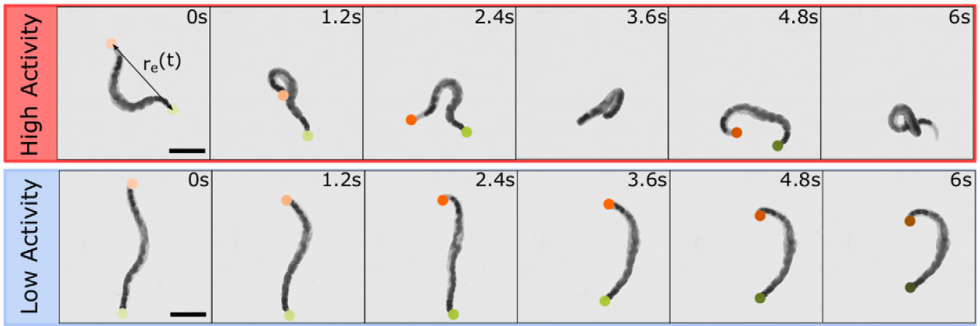
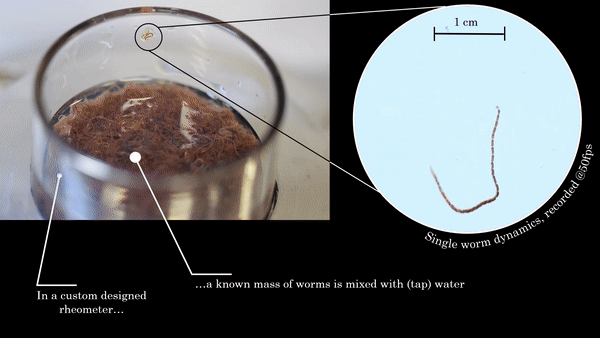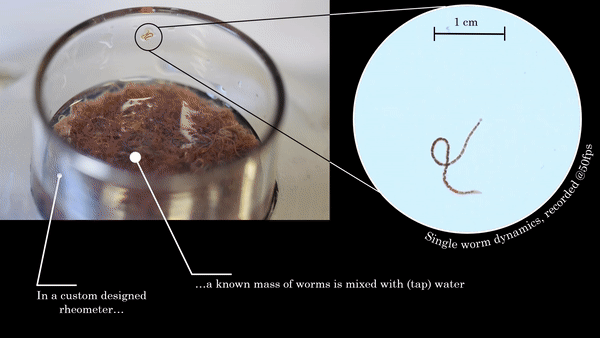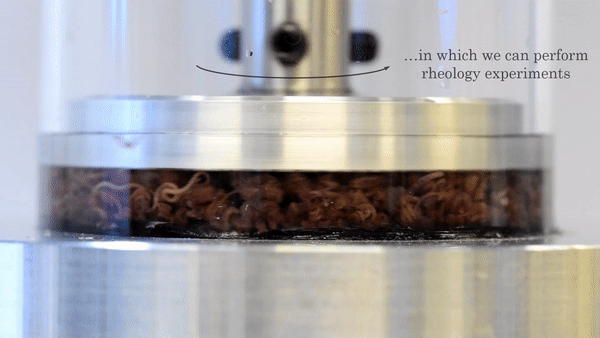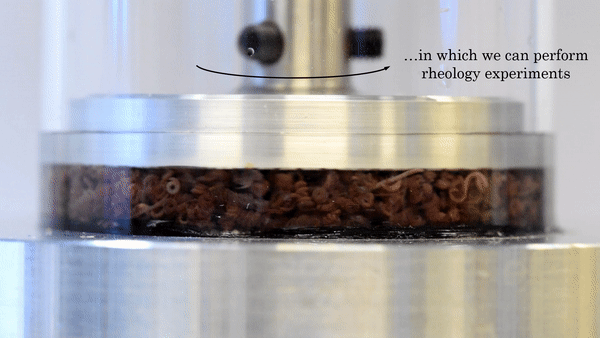
In this study, the researchers asked: how does shear thinning behavior change if the polymers in question were alive? To answer this question, they set out to measure the shear thinning properties of a mixture of worms at various levels of worm activity. Here, “worm activity” refers to how fast the worm is wriggling, which is calculated by measuring how quickly the distance between the two ends of a given worm changes. This leads to two logistical questions: how is the level of worm activity being modified, and how is the viscosity being measured?
Figure 1. This movie shows two worms, one in water (left) and one in a water + alcohol mixture (right). The worm on the right shows a decrease in activity when they are exposed to alcohol, which is one of the two ways the researchers modified worm activity in this study. Video taken from the original article.
The answer to the first question should be familiar to many humans. To make the worms less active, they were put into a solution containing water and a small amount of ethanol, the same type of alcohol found in beer, wine, and spirits. Once the worms were nice and drunk, the researchers noticed that they squirmed about more slowly, as shown in Figure 1. Thankfully, when the ethanol was removed, the worms returned to their previous level of activity! To make sure the alcohol wasn’t doing anything funny to the worms, they found a second way to reduce the activity — by reducing the temperature of the worm solution. Colder temperatures made for more chilled out worms, no pun intended.
Figure 2. This movie shows the functioning of the rheometer. The worms are placed inside a chamber between two plates. The top plate rotates with respect to the bottom plate, and the response of the material is measured. Video taken from the original article.
The researchers used a device called a parallel-plate rheometer to understand the shear thinning behavior of this living polymer system. As seen in Figure 2, a parallel-plate rheometer sandwiches a sample in between two flat plates and viscosity is measured by determining how much force is necessary to rotate the top plate, effectively pushing the material by twisting its surface. The viscosity of the worm mixtures was first determined at three different temperatures, and for worms drunk on ethanol. The results were surprising! The rheological behaviour of the low-activity worm mixtures matched with theories of polymer shear thinning quite well. It seems the worms have the same alignment properties as passive polymer solutions under shear!
So what happens when the worms are sober, more active, and wriggling around? They saw that the required twisting rate needed to thin the mixture decreased. In this case, the worm activity allowed for easier and quicker rearrangement while the mixture was pulled apart by the rheometer’s twisting motion. One can imagine that instead of needing to pull all the worms to the point of alignment, it may have been enough to nudge them in that direction and their wriggling did the rest. We can now imagine that the same thing might be true for non-living polymers: if a polymer material with shear thinning behavior is given an extra source of activity, then its thinning behavior may become more significant.
The lesson to be learned here is partly about worms, polymers, and the adverse effects of ethanol, but really this experiment is a testament to the power and generality of physical descriptions. This study teaches us about the possible behavior of an active polymer system with processes that are relevant on the scale of a few micrometers, by studying real life worms that you can see with the naked eye! In general, it is usually possible to find analog systems that have the desired properties for your study, but which are easier to manipulate. Physics then gives you the bridge between the system of interest and your simpler analog, allowing you to harness the power of interdisciplinary science to ask questions previously unanswerable.
Featured image for the article is taken from the original article.
An alien spaceship commander was preparing to drop a cone-shaped spy shuttle into the sand of a Florida beach near Cape Canaveral. The shuttle needed to burrow deep enough that any passing humans wouldn’t see it while the aliens used it to spy on Earth’s space program. “From how high should I drop the shuttle so that it is hidden?” the commander asked their science advisor. The science advisor pulled out their alien high school mechanics book, hoping to calculate this based on the laws of motion and Earth’s gravitational force.
Not so fast, alien science advisor! While the mechanics of a falling shuttle are relatively simple, the forces the shuttle would experience while penetrating the sand are much more complicated. Sand and other granular materials are composed of millions of individual solid particles that, together, may be stiff like a solid or flow like a fluid. This interstellar scientist first needed to know how sand particles interact with one another and how the uneven distribution of forces between them before dropping the cone-shaped probe.
In “Collisional model of energy dissipation in three-dimensional granular impact”, C. S. Bester and R. P. Behringer asked a similar question. In their study, they looked at the forces that a conical object (such as the alien commander’s spy shuttle) experienced as it penetrated a granular material and investigated how these forces affect the depth a conical object will burrow.
Bester and Behringer dropped conical intruders into a container filled with sand with a thin rod attached to the top of the intruder for tracking. They filmed each falling intruder from the side with a high speed camera, from which they determined the depth z, velocity, and acceleration as the intruder penetrated the sand. For a video of the experiment, see here. They used the seven intruders shown in Figure 1 . The intruders all had the same mass m but different shapes. The sharpness of the intruder’s cone-shaped tip was characterized by a parameter $latex s= \frac{2 L_{tip} }{w}$, the ratio of its length to half its width. A higher value of s corresponded to a sharper cone. They were dropped from a range of heights between 6 cm and 2 m, which resulted in intruders reaching different speeds upon impact with the sand.
Figure 1: intruders of equal mass used in the experiment, from the bluntest (s = 0) to the sharpest (s = 2.1). Image adapted from original article.
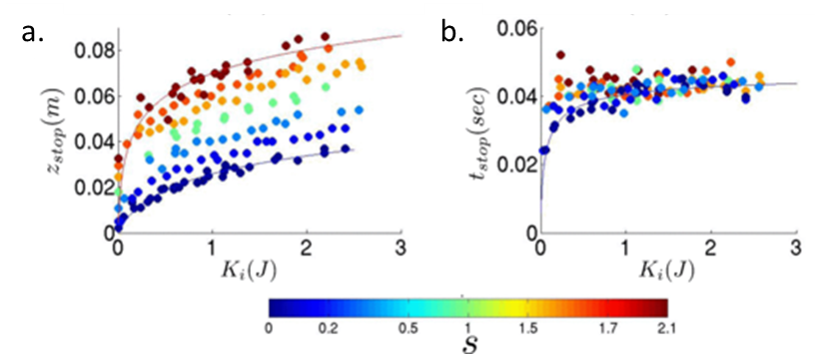
Bester and Behringer measured the stopping depth zstop and the time to stop tstop as a function of the initial kinetic energy each intruder had upon hitting the sand, $latex K_i = \frac{1} {2}m z_i$. They found that sharp intruders penetrated deeper into the sand than blunt intruders with the same kinetic energy Ki, as shown in Figure 2a. Figure 2b shows that, above an initial kinetic energy of 1 J, the time the intruders took to stop was the same regardless of shape or initial energy.
Figure 2: (a) Stopping depth as a function of kinetic energy. (b) Stopping time as a function of kinetic energy. Blue represents blunt intruders while red represents sharp intruders. Figure adapted from original article.
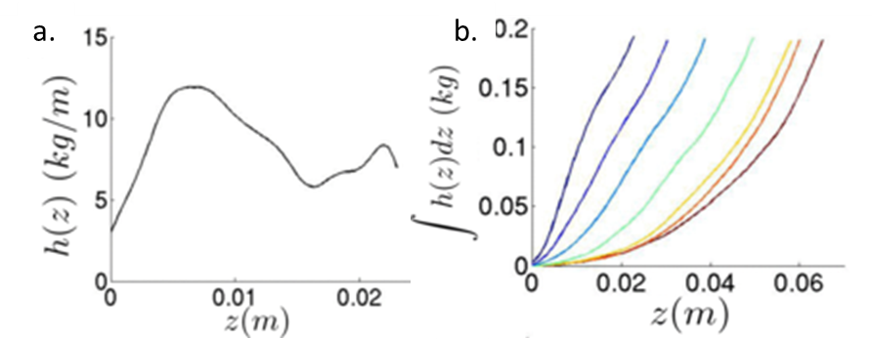
To understand what forces the intruder experiences as it comes to a stop, the authors focused on the inertial drag, or the drag caused by the pressure of the sand on the intruder. Previous studies hypothesized that the inertial drag depended on the penetration depth and was proportional to the velocity squared of the intruder as it enters the granular material. Bester and Behringer found that this was not the whole story. They calculated the inertial drag coefficient h(z) from the intruder trajectories, as shown in Figure 3a. Surprisingly, they found that the drag coefficient oscillated as the intruder penetrated the material. This suggested that the inertial drag was caused by collisions of the intruder with particles that are part of “force chains”. Force chains in a granular material are made up of connected particles that bear the majority of the forces in the material (see this earlier Softbites post for a detailed description). When the intruder hit a force chain, the drag increased due to the added resistance. The drag then decreased again when the chain was broken.
To investigate how the drag force was affected by the shape of the intruder, Bester and Behringer used the sum of the drag coefficient as the intruder penetrated the sand $latex \int {h(z) dz}$ [1]. Blunt intruders had a drag that increased nearly linearly with depth, while the dependence of drag on depth was much more curved for sharp intruders, as seen in Figure 3b. The authors suggested that the nonlinear drag for sharper cones was caused by the changing surface area interacting with the grains. Upon impact, a sharp cone only interacted with the sand through the tip. As it sunk, the area that was in contact with the sand increased nonlinearly, which resulted in larger drag.
Figure 3: (a) Drag coefficient h as a function of height z for an intruder shows fluctuations. (b) Drag dependence on depth for different intruders. The drag of blunt intruders has a roughly linear relationship with depth (blue curves) while that of sharp intruders has a nonlinear relationship with depth (yellow and red curves). Figure adapted from original article.
Bester and Behringer’s investigation into how the shape of a conical intruder falling into sand affects the forces it experiences is a beautiful example of how complex the interactions of everyday materials can be. According to their work, the aliens in our introduction should drop a pointy probe from very high up to make sure it gets buried — and also put some sensors on their probe to measure how its descent is interrupted by the force chains in the sand. The aliens may have imaginary science fiction technology that allows them to traverse light years, but even they may marvel at the countless collisions that affect the path of something they drop on the beach once they reach the Earth.
[1] The sum of the drag coefficient ($latex \int {h(z) dz}$) was calculated from the measured kinetic energy, and then the derivative of it was taken to obtain the drag coefficient h. Taking the derivative amplified the noise in the measurement. Bester and Behringer compared the sum $latex \int {h(z) dz}$. for different intruders to avoid this amplified noise.
Polymers are made of long molecules (polymer chains) consisting of shorter, repeating units called monomers. Like cooked spaghetti noodles, many polymer chains coexist in the same shared space and when too many of them overlap entanglement may occur (Figure 1). Such entangled messes of polymer chains are stiff and hard to deform, limiting the elasticity of polymer-based synthetic materials. One way of softening materials is by disentangling the polymer chains via soaking the polymer chains in a solvent, such as water. The solvent molecules in hydrogels occupy space between polymer chains driving the chains away from each other, similar to how pouring water overcooked spaghetti drives the noodles apart. This led to the discovery of hydrogels, the primary component of soft contact lenses and tissue implants [1]. But if you’ve ever worn soft contact lenses, you may know that they dry out and harden if they are not stored in a solution. This pervasive issue of hydrogel materials occurs when the solvent leaks or evaporates, affecting their mechanical properties. In this week’s post, polymer scientists develop super-soft dry elastomers (very elastic or rubbery polymers) that surpass the softness and elasticity of hydrogels, all without getting their hands wet.
Figure 1. Spaghetti pomodoro e basilico. The noodles demonstrate how long, flexible objects intertwine with each other to form an entangled complex, resembling polymer networks in hydrogels. Image courtesy of Wikipedia.
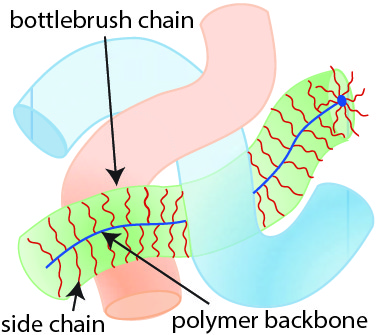
What does it take to design a polymer material that intrinsically avoids entanglement without using a large amount of solvent? William Daniel and his colleagues tackle this issue by designing a polymer chain geometry resembling a bottlebrush (shown in Figure 2). The bottlebrush geometry consists of a linear polymer backbone onto which short side chains, called bristles, are grafted. These bottlebrush-shaped chains are soft even in the absence of solvent. Instead of relying on small solvent molecules, bottlebrush networks use their bristles to keep polymer chains away from each other because bristles are too short to participate in the entanglement. As a result, the bottlebrush chains have an overall repulsion effect that resembles the repulsion effect of solvent molecules in hydrogels. Bristle repulsion allows bottlebrush polymers to surpass the elasticity of hydrogels! As an example, Figure 3 shows a compression test where a bottlebrush elastomer (on the right) retains its structure whereas a hydrogel material (on the left) fractures when compressed. Despite their similar elastic moduli, the bottlebrush elastomer displayed much greater compressibility than the hydrogel.
Figure 2. Schematic of three bottlebrush polymer chains. Each bottlebrush chain consists of a polymer backbone (linear chain) onto which short side chains (bristles) are attached. Figure adapted from the original article.
But what is an elastic modulus and why does it matter? The elastic modulus is a measure of the stiffness of a material and is given by the ratio of stress, the force causing deformation per area, to strain, the relative length by which the material is deformed by the stress. In these terms, a small modulus corresponds to low force per area resulting in significant deformation during compression – exactly what we expect of soft materials! As the entanglement density increases, a polymer chain network becomes more crowded resulting in a stiffer material [2]. Since bottlebrush bristles are too short to entangle, increasing the bristle density further reduces the entanglement density. Thus the elastic modulus of bottlebrush elastomers can be tuned by controlling the number of bristles grafted onto the polymer backbone. These bristles comprise the majority of the mass of the elastomer, e.g. 87% of the mass of the elastomer described in Figure 3.
Figure 3. Compression test of a hydrogel versus a bottlebrush elastomer. Left: PAM or poly(acrylamide) hydrogel made of 10% by weight polymer chains and 90% solvent. Right: bottlebrush elastomer made of 8% by weight backbone chains, 87% bristles and 5% solvent. Both materials have similar moduli (~2000 Pa). During compression, the bottlebrush elastomer kept its form while the hydrogel fractured. Figure adapted from the original article.
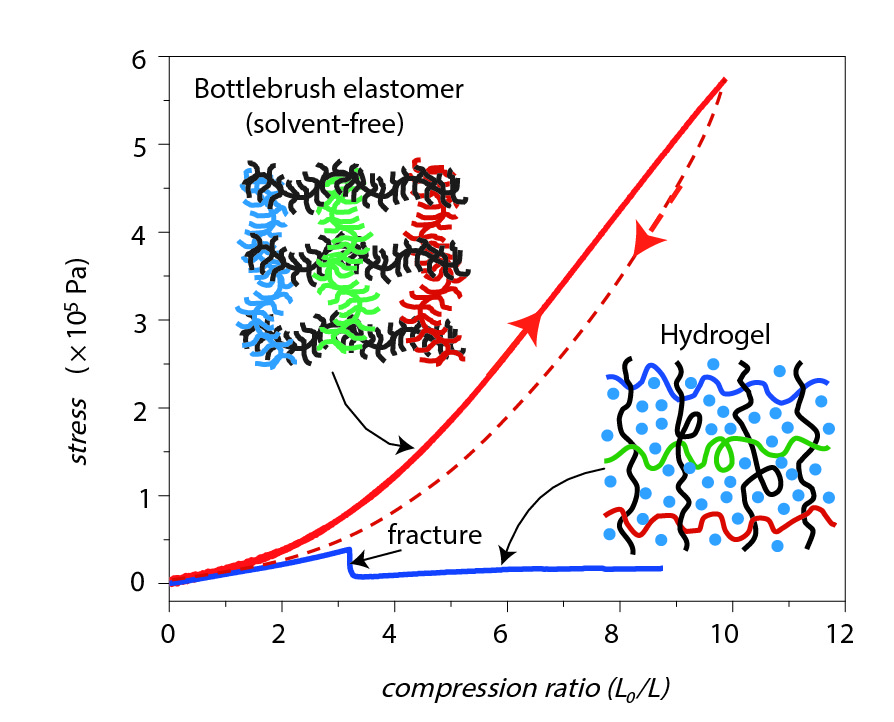
In addition to super-softness, bottlebrush networks are also highly compressible. The stress measurement in Figure 4 shows that bottlebrush elastomers (red curve) tolerated five times more stress before fracture compared to hydrogels (blue curve). Furthermore, the compression ratio (equilibrium length to compression length) of bottlebrushes was three times higher before fracturing. This means that bottlebrush elastomers are capable of sustaining much more deformation, and hence strain, than hydrogels.
Figure 4. Stress measured during compression of a bottlebrush elastomer (red curve) and a hydrogel (blue curve). The bottlebrush elastomer achieved a compression ratio (equilibrium length to compressed length) of about 10 while the hydrogel fractured at a compression ratio equal to 3. Image adapted from the original paper.
The idea of attaching bristles onto a polymer backbone in high density gave William Daniel and his colleagues control over the stiffness due to entanglement. This work expands scientists’ understanding of material properties consisting of branched polymer chains and points to a new frontier of dry supersoft materials. These new materials could play an important role in the development of soft robotics and synthetic biological tissues.
[2] In polymers, the entanglement contribution to the elastic modulus is given by the modulus equation:
Ge = neRT,
where ne is the number of chains involved in entanglement per unit volume, R is the universal gas constant, and T is the temperature. The modulus equation suggests that polymer materials become stiffer when heated. As temperature increases, the polymer network gains kinetic energy to attain structures with more “randomness”, or entropy. This increased randomness in polymer networks leads to more entangled states (higher entropy) that makes the material less prone to lengthening, hence more resistant to deformation.
In these unprecedented and fluid times, conferences and symposia have gone virtual as STEM collectively settles into a new normal. Many large meetings, like the formerly “in-person only” American Physical Society (APS) and American Chemical Society (ACS) national meetings, have been canceled or transitioned to virtual-only participation this year. The 2021 Spring APS meeting will go virtual as well. I love big in-person meetings and have shied away from virtual alternatives thinking they would not provide the same feeling of community with my fellow scientists. However, the isolation of quarantine and the desire to get comfortable with the “new normal” motivated me to step out of my comfort zone and into the world of virtual science meetings this summer. So, when the opportunity to attend the 2020 Virtual Polymer Physics Symposium (VPPS) arose in July, I jumped at the chance to participate.
The Polymer Physics GRS is held biennially; the last seminar was in 2018 and, due to the pandemic, the next one won’t occur until 2022. “Two years feels like a lifetime to ECRs and we know many of our colleagues will be at different institutions and career stages in 2022, so we wanted to create a space for them to share and discuss their recent research,” explains Konane. The 2020 VPPS connected more than 100 scientists, including 20 ECRs presenting their work across four oral presentation sessions during the two-day event. In the spirit of the Polymer Physics GRS, this new virtual event also incorporated professional development and discussion sessions, including a Mentorship Panel and the “Dispersity and Diversity Hour” discussion focused on how to increase diversity, equity, and inclusion (DEI) in the field of polymer physics and the broader scientific community.
The Mentorship Panel included researchers at different career stages working in academia, industry, or government research. The discussion was focused on steps students and postdocs can take to prepare for future careers in polymer physics as we adapt to a global pandemic. The panelists emphasized the quarantine-proof nature of computational work since it can often be done anywhere including at home and encouraged experimentalists to broaden their computational skill set. Both Nate Lynd, assistant professor at UT Austin, and Michelle Sing, an engineer at Braksem USA, suggested that experimentalists should become familiar with Python as a first step. Debra Audus, a scientist at NIST, highlighted lab work automation and strategic experiment planning to maximize “in-lab” time during the transition to shift-style lab work that many American universities have adopted as a way to overcome the challenges related to performing socially distanced science.
Participating in the “Dispersity and Diversity Hour” required some homework. The organizers asked all attendees to prepare for the discussion by reading about the experience of Black researchers in STEM (links below). Ben Yavitt, a Stony Brook University postdoc, opened the event by emphasizing that the goal of the discussion was to brainstorm potential solutions to address the issues spotlighted by national movements such as #BLM, #ShutdownSTEM, and #BlackintheIvory. More than 70 attendees participated in small group discussions across 15 breakout rooms led by volunteer discussion leaders. This event was an important first step in raising awareness of the necessary academic culture shift required to empower more scientists of color to pursue careers in soft matter and polymer physics.
Overall, the meeting events were very well moderated and designed for maximum virtual engagement during both the science presentations and the discussion sessions. For me, attending this event clarified the current trajectory of the polymer physics field as it transitions from fundamental studies to applied research focused on exploiting polymer physics for advanced technology. This transition in research focus was evident in some of the science presented during the symposium. While I was apprehensive about the shift to virtual meetings and conferences, attending the 2020 VPPS has won me over. I highly recommend taking advantage of the proliferation of virtual science conferences and symposia to stay engaged, learn about new science and perspectives, and do some networking along the way!
Disclosure: I am acquainted with Konane Bay, one of the event co-organizers. She was a graduate student in my department, UMass Amherst Department of Polymer Science and Engineering. However, I was not involved in any way with the organization of the VPPS.
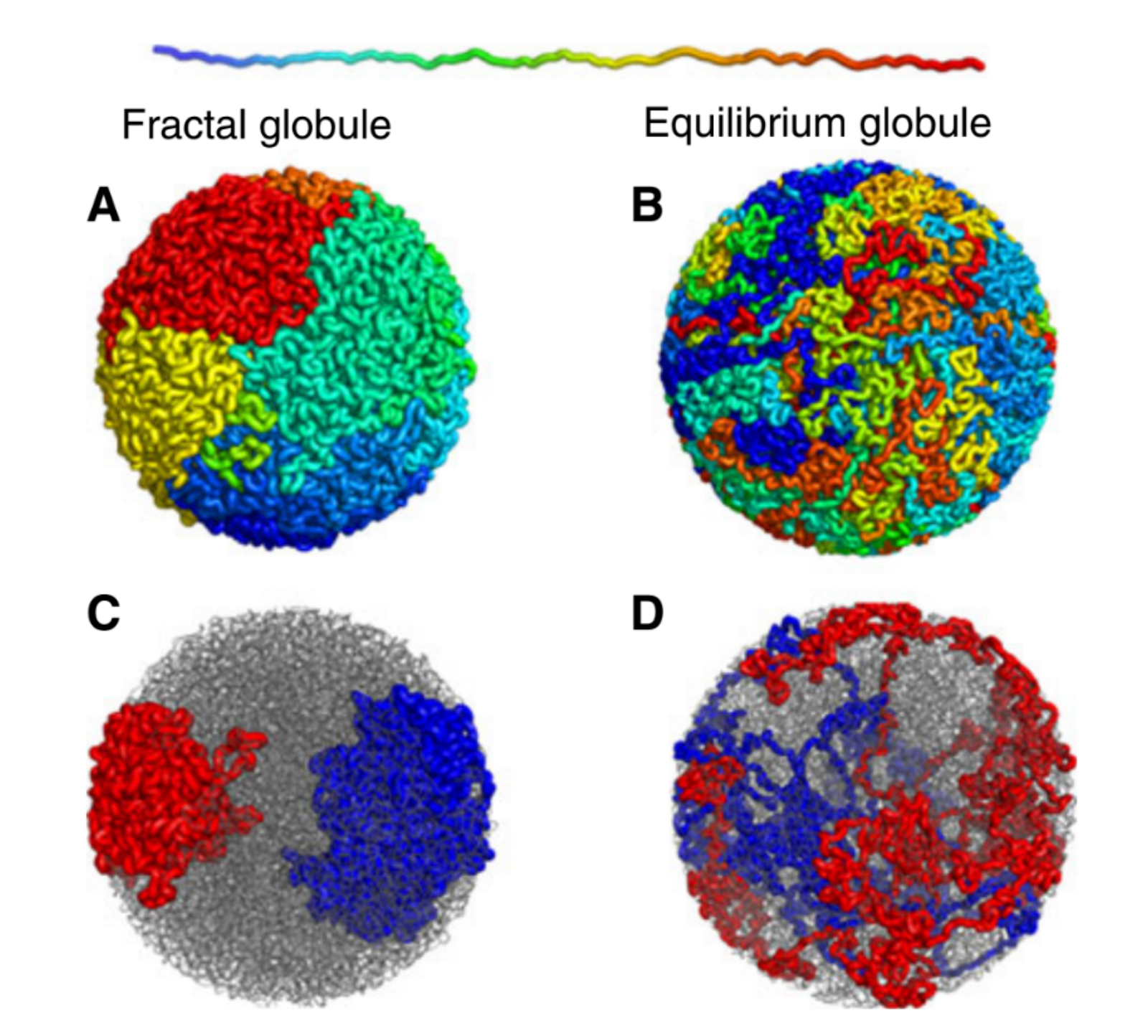
The entirety of our genetic information is encoded in our DNA. In our cells, it wraps together with proteins to form a flexible fiber about 2 metres long known as chromatin. Despite its length, each cell in our body keeps a copy of our chromatin in its nucleus, which is only about 10 microns across. For scale, if the nucleus was the size of a basketball, its chromatin would be about 90 miles long. How can it all fit in there? To make matters worse, the cell needs chromatin to be easily accessible for reading and copying, so it can’t be all tangled up. It’s not surprising then that scientists have been puzzled as to how this packing problem can be reliably solved in every cell. The solution is to pack the chromatin in a specific way, and research suggests that this may be in the form of a “fractal globule”.
An equilibrium globule is the state that a polymer (a long repetitive molecular sequence, like chromatin) takes when it is left for a long time in a liquid that doesn’t dissolve it well. In such a liquid, the polymer is more attracted to itself than the molecules around it, so it collapses into a globule to minimize the amount of contact between itself and its surroundings. The resulting object is much denser than typical polymers in good solvents and is dense enough to fit inside a nucleus. However, like stuffing headphone cables into your pocket, it develops many knots and its different regions mix with one another.
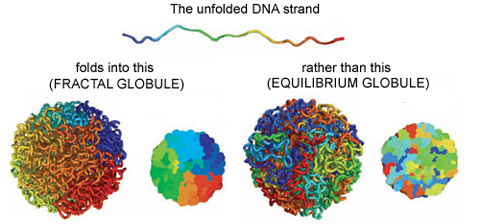
On the other hand, if you change the polymer’s environment fast enough that it doesn’t have the time to fully equilibrate, then every piece of the polymer will locally collapse into its own globule. In other words, the polymer forms a globule made of smaller globules and is called a fractal globule. Fractals are objects which look the same at all scales, like the edge of a cloud or the coastline of England. If you zoom in or out on either of these objects, they look more or less the same. This isn’t an “equilibrium” state, meaning it will slowly fall out of this configuration. However, until the whole polymer equilibrates (which takes a long time), the chain has many desirable properties.
Figure 1. Simulated examples of fractal (A,C) and equilibrium globules (B,D), showing compartmentalization of different portions of the polymer. The chain color goes from red to blue as shown above. Compartmentalization means that parts of the chromatin stay near other parts with the same color (adapted from paper [1]).
We are interested in these globule states because they are dense enough that a globule of chromatin can fit inside of a cell nucleus. But it’s not enough to simply fit inside; the cell needs chromatin to avoid forming knots, since getting tangled would prevent the cell from properly reading its own DNA. Live cells also keep their chromatin nicely compartmentalized, that is, different regions along the genome stay spatially separated from one another. Unlike equilibrium globules, fractal globules have few knots and are also compartmentalized! To get a better picture for what this means, Leonid Mirny performed simulations of the different types of globules. Figure 1 shows the results of these simulations, highlighting how different the two states look in terms of knotting and separation of regions of the polymer.
So it seems that the fractal globule state has all the properties we need for a good model of chromatin! But, as scientists, we know that no matter how well a theory fits the characteristics we want it to have, we need experimental evidence before believing anything. In the case of this fractal globule model for genome organization, evidence has come in the form of “contact probability maps”. These are collected from large populations of cells whose DNA is cut, spliced, and read in such a way that allows for a measurement of the probability that any two sites on the chromatin are touching at any given time. Among other things, these maps give us information about how chromatin is packed. So the question becomes, “what does the fractal globule model predict a contact probability map to look like?”
The fractal globule model doesn’t make exact predictions about where one will find specific segments of chromatin, but it does predict a contact probability as a function of distance between two sites, s. Specifically, the model predicts that the contact probability between two sites scales like 1/s. Meaning, if I look at sites that are twice as far apart along the polymer, then they are half as likely to be touching. This 1/s scaling is what was observed on intermediate scales (about 100,000 to 6 million base pairs) by looking at contact probability maps averaged over a whole population of cells.
We still don’t know how the cell maintains and tunes this fractal globule state, and we still have not developed a dynamic version of this picture, which is necessary since it is well-established that the chromatin in our cells is far from static. But this study gives us a new picture of how chromatin is organized inside cells. It isn’t randomly configured like headphone cables in your pocket or a ball of yarn. Rather it is folded onto itself in a self-similar way. This model is attractively simple, requires little fine-tuning, all while producing a long-lived state with segregated territories and easily accessible genes.
[1] Mirny, Leonid (2011), The fractal globule as a model of chromatin architecture in the cell. Chromosome Res.
Featured image for the article is taken from Wikimedia Commons.
I was ready. I was so ready. I had all my chargers and AV adapters. My presentation was backed up on a USB drive. I had every talk I wanted to go to on my calendar. I had sent emails to professors I wanted to meet and network with. I reached out to friends I only see in March in a different city every year. It was 10 pm on Saturday, February 29. My flight to Denver was leaving at 6 am in the morning. Then the email arrived —
URGENT: 2020 APS March Meeting in Denver, CO – CANCELED. Due to rapidly escalating health concerns relating to the spread of the coronavirus disease (COVID-19), the 2020 APS March Meeting in Denver, CO, has been canceled. Please do not travel to Denver to attend the March Meeting. More information will follow shortly.
That was it. All the preparation, all the planning, thrown out the window because of the novel coronavirus that was quickly spreading throughout the world. Writing this more than 2 weeks later in the face of a bona fide pandemic with entire countries locked down, the decision was extremely prescient and prudent. At the time, it felt a bit like being forced to eat vegetables as a child.
But as soon as the APS March Meeting was cancelled, would-be participants started self-organizing their own virtual conferences. Fittingly, the Division of Soft Matter (DSOFT), which largely studies self-organization, led the charge. Professor Karen Daniels from NC State University used Twitter to announce that plans were underway to host virtual talks, and one day later had a place for people to sign up to host and give the talks they had prepared for Denver. The Division of Biological Physics (DBIO) also followed suit, led by Professor Phillip Nelson from the University of Pennsylvania. By the end of the week, a remarkable 60 sessions between both DSOFT and DBIO were held online, all due to grassroots organization over social media and word-of-mouth.
The method of choice was definitely Zoom, with BlueJeans and Google Hangouts also being used. The first virtual session I attended was (appropriately) The Physics of Social Interactions, hosted by Professor Orit Peleg. I was shocked to see over 50 people in the Zoom meeting, but even more shocked at how smoothly the entire process unfolded. Professor Peleg kept the speakers on time and the “Raise hand” feature on Zoom made asking and answering questions easy and painless. The first virtual DBIO session held was the Delbruck Prize session, where Professor Jim Collins from MIT was being honored for his work in synthetic biology, including some pertinent work on cheap and fast testing for various diseases. That session had over 100 participants and still went just as smoothly as the smaller ones. I was also able to give my talk virtually to a total of 5 people present, but I’ll put my ego aside to be happy I gave it at all.
These virtual talks definitely do not replicate the conference going experience, but they do come close. In my experience, many people attending talks are already on their laptops attending to their own business, so listening to a virtual talk while answering emails from the comfort of one’s desk felt familiar. When it comes to gathering scientific information, the virtual meeting was a great success. Even those who were unable to attend the meeting were able to see some talks of interest to them. However, as one might expect, the social component of the meeting was largely lost. Hallways are where the magic really happens at conferences, and the closest approximation the virtual APS meeting had was Twitter. Nevertheless, sending a quick email to a speaker whose talk you liked could accomplish most of the networking one might try to accomplish.
While it’s hard to find a silver lining in COVID-19, these virtual meetings will almost certainly open the doors for future scientific meetings allowing virtual talks to be given by those who cannot attend. As scientists, we should model what it looks like to lower our carbon footprint without impeding professional advancement. In addition, virtual talks give access to meetings for those who cannot physically travel for financial, familial, and/or physical reasons, promoting the inclusion of traditionally marginalized populations in the larger scientific community. As more meetings get corona-cancelled, such as the upcoming APS April Meeting, the use of virtual conferences will only become more streamlined and normalized. I personally believe this will change the way scientists communicate around the world for the better, even if it took a pandemic to get here.