Original paper: Motility-induced fracture reveals a ductile-to-brittle crossover in a simple animal’s epithelia
Content review: Heather Hamilton
Style review: Pierre Lehéricey
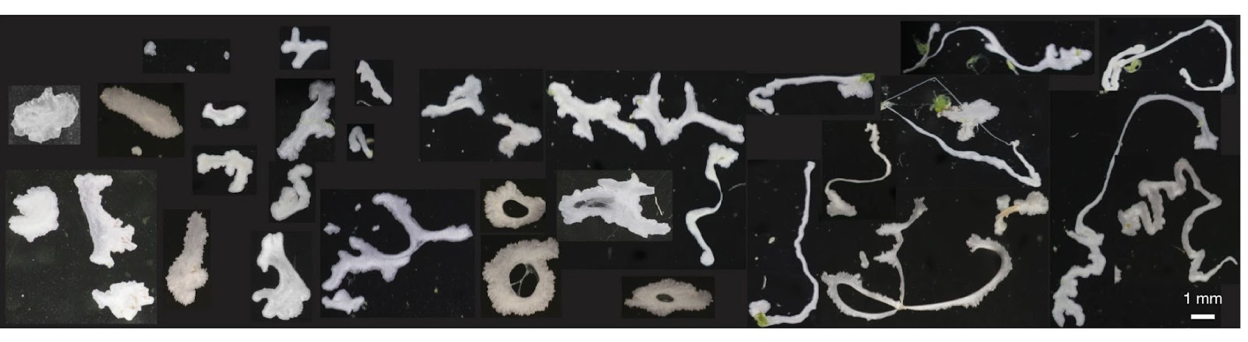
Meet Trichoplax adhaerens, a microscopic marine animal from one of the oldest known branches of the evolutionary tree. It looks like a microscopic cell sandwich: two layers of epithelial cells (which make up the surfaces of our organs), with a layer of fibre cells in between. As depicted in Figure 1, T. adhaerens takes a wide variety of shapes from disks to loops to noodles and more. Oddly, T. adhaerens ruptures when it moves around, a self-induced fracture behavior that has recently captured the attention of physicists and engineers. Fracture is the technical term describing the process by which an object breaks into distinct pieces due to stress. These animals push their epithelial tissue to the breaking point, forming incredible and extreme shapes before separating altogether. This is a surprising behavior for epithelia, which usually prefer to maintain their integrity. By modeling how T. adhaerens rips itself apart when moving, we can improve our understanding of how soft materials and especially biological tissues behave on the verge of breaking.
Prakash, Bull, and Prakash conducted a two-pronged analysis of fracture in T. adhaerens: live imaging to record the fracturing in real time and computational modeling to simulate the response of the tissue when stretched too far. The drastic mechanical behavior in question also motivated the researchers to perform a more general inquiry into the competition between flow and fracture in materials that are dramatically deformed relatively quickly. Flow is like stretching out a piece of chewing gum, whereas fracture is like snapping the gum in two. The computational model proposed by the authors helped paint a clearer picture of what happens when T. adhaerens rips apart.
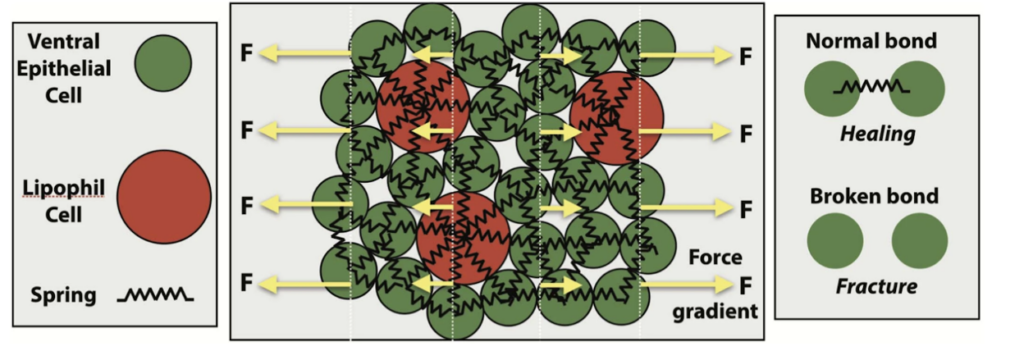
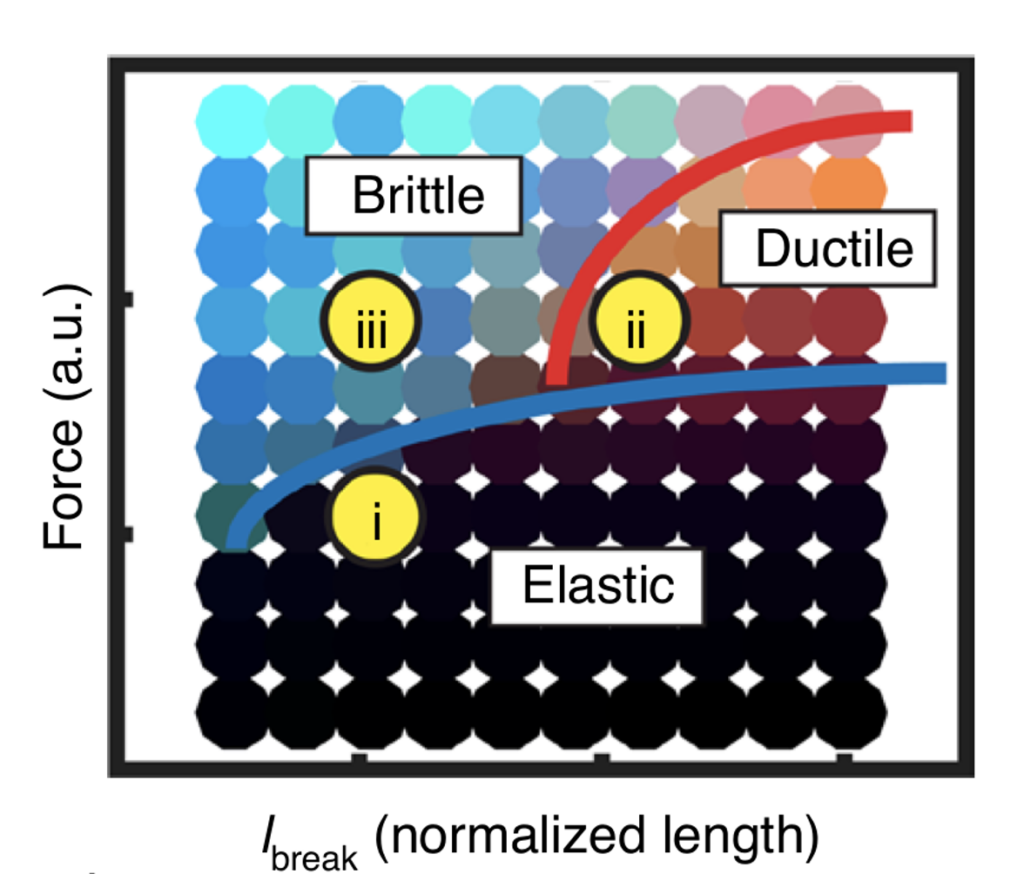
The computational model that the researchers used is based on a sticky ball and spring model, as shown in Figure 2, where each ball represents a cell and each spring represents the sticky junctions that cells use to adhere to one another. The springs break if the balls move too far away from each other, which represents cells being unstuck from their neighbors. Two cell types are represented in the model epithelial layer in Figure 2: epithelial cells, which are small and comprise the bulk of the tissue, and lipophil cells, which are larger and less common. Using this model for living tissue, the authors conducted computational simulations where the tissue was stretched to a breaking point. They found that there are three possible tissue behaviors that depend on the strength of the driving force applied to the simulated tissue. For weak forcing (low stress), the tissue behaved elastically and so responded in such a way that it could recover its original shape. For intermediate forcing (medium stress), the tissue underwent a “yielding transition” where the material transitioned from elastic response to plastic response. During plastic response, permanent distortions occurred in the material, and the material could not recover its original shape. In this case, the tissue is ductile and undergoes local changes, like cells interchanging with neighboring cells, to relax some of the pent-up stress. For stronger forcing (high stress), the tissue undergoes brittle fracture where the bonds between cells break with little opportunity for relaxation. The three behaviors in the model represent a transition from elastic to ductile to brittle responses. Using this model of tissue response to applied force, the authors mapped the conditions that lead to different tissue behaviors, as sketched in Figure 3.
Guided by a better understanding of tissue mechanics thanks to the computer model, the authors experimentally measured the brittle and ductile responses in T. adhaerens. They found that both material responses can occur in our microscopic friend. The ability to access both regimes is important because the ductile response yields by flowing (helping form the longer shapes in T. adhaerens) whereas the fracture response accounts for asexual reproduction by splitting into two separate new individuals. The authors’ combined approach of experimental data that motivated the development of a computer model, which in turn guided further experimental inquiry, is an important modern scientific paradigm. Both approaches are incredibly important tools in the biological and soft matter sciences’ toolkit. Joint application of these tools lets us draw general conclusions from specific experiments as well as apply those general conclusions back to answer specific questions – like explaining how T. adhaerens achieves the diversity of shapes in Figure 1 and how this relates to its hardiness and evolutionary goal of reproduction. Further, the epithelial layer computational modeling technique generalizes this tissue mechanics study to help us describe fracture versus flow in any living tissue, including our own.