

![Berginski M, Vitriol E, Hahn K, Gomez S [CC BY 2.5 (https://creativecommons.org/licenses/by/2.5)], via Wikimedia Commons](https://softbites.org/wp-content/uploads/2019/02/high-resolution-quantification-of-focal-adhesion-spatiotemporal-dynamics-in-living-cells-pone.0022025.s014.ogv.jpg)
Original paper: Measuring mechanical tension across vinculin reveals regulation of focal adhesion dynamics
“USE YOUR LEGS!” That’s what might have been yelled at you the first time you went climbing. We are so used to walking or running that we don’t even think about how we do it. But when we face a new environment, such as a steep slope, we realize that finding the best strategy to move through space is not so easy. Now, imagine you are as small as few dozens of microns, without legs or arms, and you live in a viscous fluid. How would you move? This is the question biologists who are interested in cell movements have been trying to solve. By observing cells under a microscope, they saw that depending on their type or their environment, cells exhibit a wide variety of motion strategies. However, one thing never changes: cells need to exert forces on their environment to move. To do so, some kinds of cells create structures called focal adhesions. These structures are made up of several proteins, assembled on the outside of the cell. Like tiny bits of double-sided tape, their purpose is to stick the cell to whatever is nearby (see Figure 1). In slightly more technical language, focal adhesions connect the molecular skeleton of the cell to a substrate.
Cells can exert forces on their environment through focal adhesions. While it is possible to measure these forces outside the cell by engineering some force-sensing substrate [1], it is much trickier to understand what happens inside the cell. Accessing these forces inside the cells is the challenge Grasshoff and colleagues tackled in their 2010 paper.
In order to measure a force, the most straightforward method is to use a spring. A spring is a stretchable object for which, after calibration, we can relate its extension to the applied force. Therefore, a force can be measured by measuring the length of the spring. To measure the forces focal adhesions apply on the cell, one would need to inject tiny springs in the cells and connect them to the exerting-force structures.
To do this, the authors had the idea of taking advantage of a silk protein, produced by a spider, which is literally a molecular spring. Thanks to genetic tools, a part of the gene of this silk protein could be inserted within a gene called vinculin. The vinculin gene produces a protein that is an essential part of the focal adhesion structure. As shown in Figure 2A, vinculin connects the protein filaments of the cell skeleton to the outside of the cell (the extracellular matrix). The researchers engineered an artificial variant of vinculin that includes a molecular spring, derived from the silk protein, right in the middle of the naturally occurring vinculin molecule (see Figure 2B).
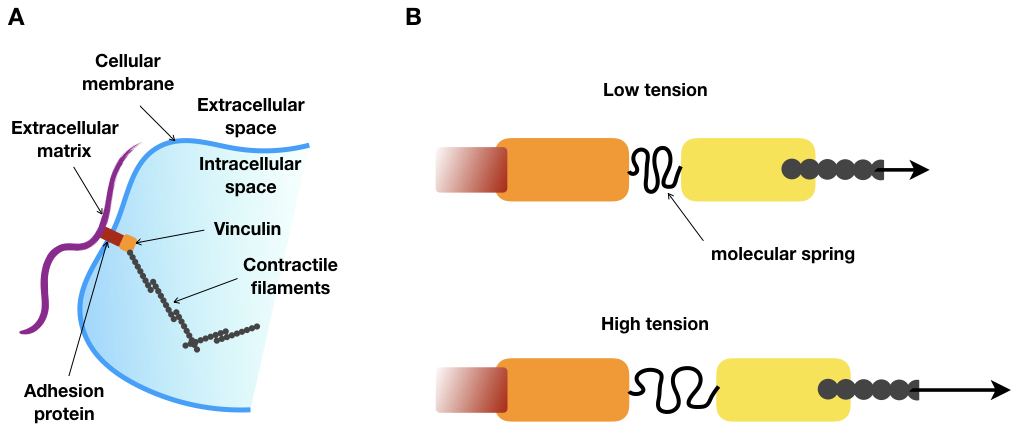
After verifying that cells that are genetically modified to include the engineered focal adhesion protein behave normally, the next step was to measure the molecular spring extension. However, measuring distances at the molecular scale is not a piece of cake. For instance, the typical extension of such a spring is 6 nanometers, which is, by far, below the resolution of the best optical microscopes [2]. To circumvent this limitation, Grasshoff and colleagues took advantage of the Förster resonance energy transfer (FRET) effect to measure the distance between the two vinculin domains. The FRET effect takes place between two fluorescent molecules very close in space. A fluorescent molecule, when excited by a light at a precise wavelength, emits a light at a longer wavelength. But if a second fluorescent molecule is close enough, the first molecule (the donor) can directly transfer its energy to the second molecule (the acceptor). Then, the acceptor will emit light at an even larger wavelength than the donor’s. Consequently, the FRET intensity can be computed by measuring the relative emissions of the donor and acceptor molecules: the closer the acceptor is to the donor, the more energy the acceptor will absorb and re-emit. Furthermore, and importantly for this application, the efficiency of this process is very sensitive to the distance between the donor and the acceptor As a result, the distance between the two molecules can be measured with great precision (sub-nanometer) by measuring the intensity of the FRET effect. Therefore, the authors further engineered the vinculin protein by placing the molecular spring between two fluorescent molecules (Figure 3, yellow and red circles) that were capable of undergoing the FRET effect to measure the extension of the molecular spring.

At this point, the authors had a method for measuring the tension intensity across vinculin molecules just by looking at the FRET intensity. In this way, they could generate a tension map across the contacts of the cell with its environment. They saw that focal adhesion under high tension leads to a growth of the size of the focal adhesion which relieves it from its high tension. Perhaps surprisingly, they also showed that regions where the contact is extending (protruding areas) are under higher tension than regions where the contact is receding (retracting areas), as shown in Figure 4.
In this paper, the authors developed a new technique to measure forces inside cells. By conducting single-molecule experiments, they even could calibrate their engineered molecular spring and relate the FRET intensity to absolute values of forces (in the order of a few piconewtons [3]), paving the way to a whole class of new FRET-based force sensors with different stiffnesses, which can now be used in other structures inside cells.
Everything started with adding a spider silk gene in a cell. Such mutant cells have the amazing power of shading light on the cellular force machinery. But “with great power, comes great responsibility” as another spider mutant has once been told.

[1] These techniques are called traction force microscopy. The deformation of calibrated substrate (either a gel or micropillars) is measured to calculate the forces exerted by the cell.
[2] Classical optical microscopes have a typical resolution of around 200 nm. New techniques of super-resolution microscopy reach a resolution of a few dozens of nanometers.
[3] To give you a sense of this order of magnitude, when you hold a pen of, let’s say 10 g, you apply a force of 0.1 N. At the cellular level, cells exert on their environment forces in the order of dozens of nanonewtons (according to this study). At the molecular level, DNA has been manipulated applying forces in the same range as the vinculin tension: 1-100 pN (according to this study).
One Reply to “The Peter Parker cell”