Original paper: Individual-based model of larval transport to coral reefs in turbulent, wave-driven flow: behavioral responses to dissolved settlement inducer
Lost, alone, and buffeted by ocean currents: this is the beginning of life for many oceanic larvae. These tiny organisms, often only 100 micrometers in diameter, must seek a suitable new habitat by searching over length scales thousands of times their own. But searching for something you can’t see while being dragged this way and that by ocean currents can’t be easy. How do these microscopic creatures make sense of the turbulent world around them and find their home?
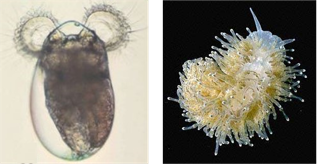
To answer this question, today’s paper studies a species of sea slug, Phestilla sibogae. These sea slugs have two forms, the baby larval form (Fig. 1 left), which travels through the ocean, and the adult sea slug form (Fig. 1 right), which lives and feeds on their coral prey. After they are born, the young larvae first swim toward light, instinctively leaving their parents’ reef. When they are old enough to settle down and become adults, they must search for a new reef to call home. The metamorphosis from larva to slug is only triggered when the larvae have settled on their coral prey.
The coral prey release a chemical that the sea slug larvae can smell. The chemical acts as an on-off switch for the larva. When there is no chemical, the larva swims in a straight path in a random direction at 170 micrometers per second. Upon encountering a strong enough chemical smell, the larva stops swimming and sinks at 130 micrometers per second. We know how the larva move but how does this movement affect how many and how quickly the larvae make it to the reef? To understand the larvae transport, we need to understand the larval environment.
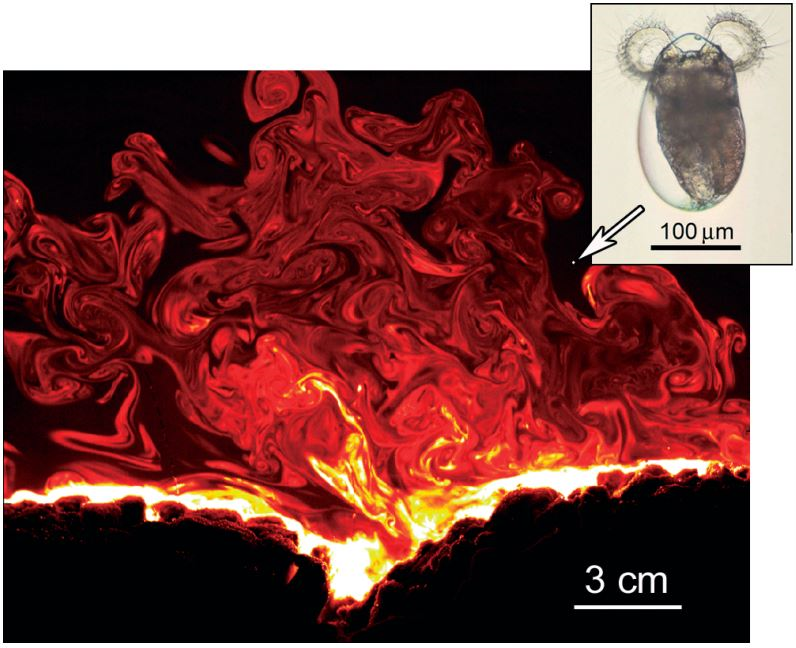
To investigate the transport of larvae to the reef, Koehl and coauthors build on previous work to create a computer simulation with both the larva swimming behavior and the larva environment based on experimental measurements. To model the background flow environment, they include the net flow, waves, and turbulence. The flow parameters are fit to experimental measurements made in wavy shallow waters in Hawaii [1]. In a similar way, the researchers use experimental measurements to model the swimming behavior of the larvae.
In their simulation, the researchers are able to alter both the environment and the larva swimming behavior and ask what, if any, advantage the on-off swimming behavior brings. The advantage is measured using the steady state larva transport rate, defined as the percentage of the larvae that make it to the reef each minute. With the steady-state approximation, the percentage of larvae that make it to the reef each minute is constant over time.
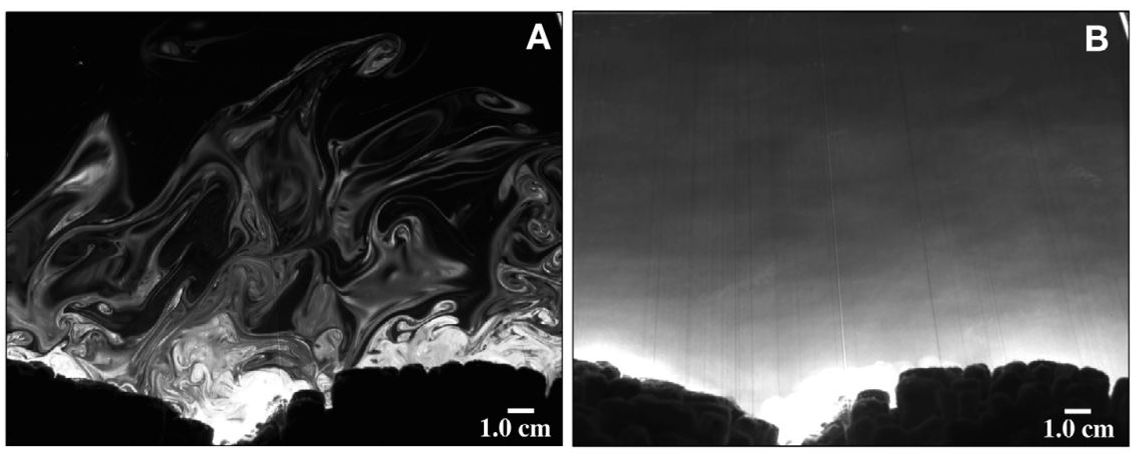
First, the researchers investigate whether or not the streakiness of the concentration pattern is an important factor in determining how many larvae reach the coral. When trying to understand how larvae reach the coral, previous researchers made the simplifying assumption that the concentration pattern of the chemical the larvae follow is smooth and uniform over time. As we saw in the streaky pattern in Figure 2, this is not a realistic assumption. But just how wrong is this it? To answer this question, the researchers compare the chemical distribution measured at a single moment in time (Fig. 3A) to the chemical distribution obtained by taking the average of the distribution measured at many different times. This averaging process produces a smoother distribution than would be seen in reality (Fig. 3B). On comparing the two different chemical distributions, the researchers find the larvae transport rates are overestimated by up to 10% in the unrealistic time-averaged environment.
Secondly, because the concentration pattern affects the transport rate, the on-off swimming behavior must affect the transport of the larvae to the reef. In their simulation, the transport rate for naturally swimming and sinking larvae is 45% per minute. The researchers test how the larva behavior affects this transport rate by separately turning off the swimming and sinking behavior of their simulated larva. If a larva sinks but does not swim, the transport rate changes to 20% per minute. If the larva swims but doesn’t sink, the transport rate changes to 25% per minute. Without their on-off switch, the larvae are reliant on the background flow or randomly swimming downwards to be transported to the coral.
From these transport rates, we can understand the relative importance of larval behavior and larval environment. For example, we now know that if the environment was no longer turbulent or if the larvae could no longer swim, the larvae’s rate of transport to the reef would change significantly. This impacts both how many larvae survive to adulthood and where in the ocean the adult sea slugs end up. Building on this work, predictions have also been made for many different species of larvae [2]. From these studies, we not only can get an idea of how local and global populations spread in their natural environments but also how a simple on-off process can help an organism to successfully navigate a complex environment.
[1] See https://academic.oup.com/icb/article/50/4/539/652640, for an overview of how researchers characterize the larva environment.
[2] See https://link.springer.com/article/10.1007%2Fs00227-015-2713-x for more details. Here, the researchers measure both the concentration of chemical and the flow above the reef simultaneously (as described in [1]). With this, they look more generally at the problem of settling on surfaces, investigating a variety of swimming properties and settling sites rather than a specific species.