Original article: Collisional model of energy dissipation in three-dimensional granular impact
An alien spaceship commander was preparing to drop a cone-shaped spy shuttle into the sand of a Florida beach near Cape Canaveral. The shuttle needed to burrow deep enough that any passing humans wouldn’t see it while the aliens used it to spy on Earth’s space program. “From how high should I drop the shuttle so that it is hidden?” the commander asked their science advisor. The science advisor pulled out their alien high school mechanics book, hoping to calculate this based on the laws of motion and Earth’s gravitational force.
Not so fast, alien science advisor! While the mechanics of a falling shuttle are relatively simple, the forces the shuttle would experience while penetrating the sand are much more complicated. Sand and other granular materials are composed of millions of individual solid particles that, together, may be stiff like a solid or flow like a fluid. This interstellar scientist first needed to know how sand particles interact with one another and how the uneven distribution of forces between them before dropping the cone-shaped probe.
In “Collisional model of energy dissipation in three-dimensional granular impact”, C. S. Bester and R. P. Behringer asked a similar question. In their study, they looked at the forces that a conical object (such as the alien commander’s spy shuttle) experienced as it penetrated a granular material and investigated how these forces affect the depth a conical object will burrow.
Bester and Behringer dropped conical intruders into a container filled with sand with a thin rod attached to the top of the intruder for tracking. They filmed each falling intruder from the side with a high speed camera, from which they determined the depth z, velocity, and acceleration as the intruder penetrated the sand. For a video of the experiment, see here. They used the seven intruders shown in Figure 1 . The intruders all had the same mass m but different shapes. The sharpness of the intruder’s cone-shaped tip was characterized by a parameter $latex s= \frac{2 L_{tip} }{w}$, the ratio of its length to half its width. A higher value of s corresponded to a sharper cone. They were dropped from a range of heights between 6 cm and 2 m, which resulted in intruders reaching different speeds upon impact with the sand.

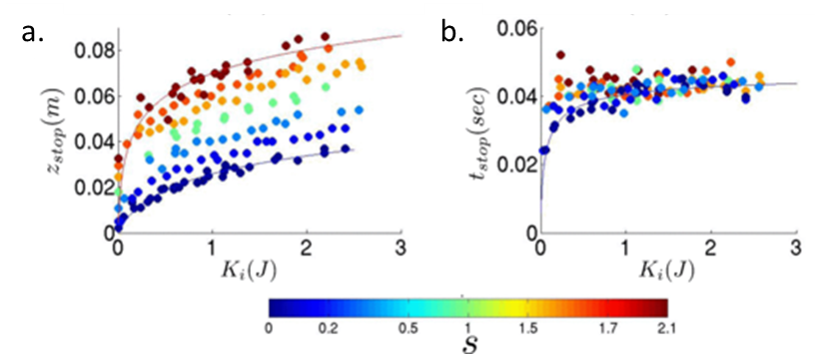
Bester and Behringer measured the stopping depth zstop and the time to stop tstop as a function of the initial kinetic energy each intruder had upon hitting the sand, $latex K_i = \frac{1} {2}m z_i$. They found that sharp intruders penetrated deeper into the sand than blunt intruders with the same kinetic energy Ki, as shown in Figure 2a. Figure 2b shows that, above an initial kinetic energy of 1 J, the time the intruders took to stop was the same regardless of shape or initial energy.
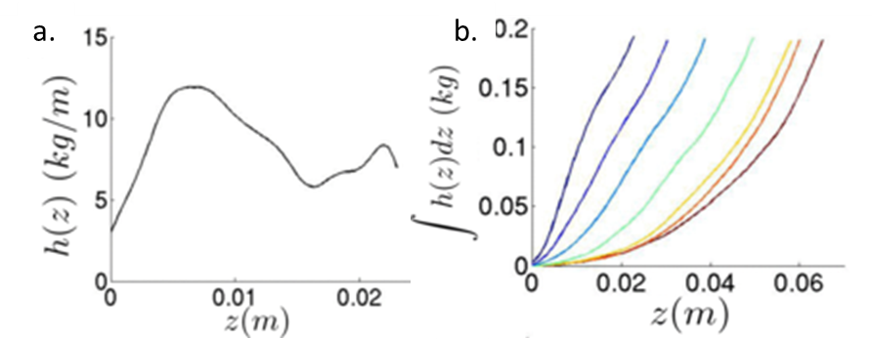
To understand what forces the intruder experiences as it comes to a stop, the authors focused on the inertial drag, or the drag caused by the pressure of the sand on the intruder. Previous studies hypothesized that the inertial drag depended on the penetration depth and was proportional to the velocity squared of the intruder as it enters the granular material. Bester and Behringer found that this was not the whole story. They calculated the inertial drag coefficient h(z) from the intruder trajectories, as shown in Figure 3a. Surprisingly, they found that the drag coefficient oscillated as the intruder penetrated the material. This suggested that the inertial drag was caused by collisions of the intruder with particles that are part of “force chains”. Force chains in a granular material are made up of connected particles that bear the majority of the forces in the material (see this earlier Softbites post for a detailed description). When the intruder hit a force chain, the drag increased due to the added resistance. The drag then decreased again when the chain was broken.
To investigate how the drag force was affected by the shape of the intruder, Bester and Behringer used the sum of the drag coefficient as the intruder penetrated the sand $latex \int {h(z) dz}$ [1]. Blunt intruders had a drag that increased nearly linearly with depth, while the dependence of drag on depth was much more curved for sharp intruders, as seen in Figure 3b. The authors suggested that the nonlinear drag for sharper cones was caused by the changing surface area interacting with the grains. Upon impact, a sharp cone only interacted with the sand through the tip. As it sunk, the area that was in contact with the sand increased nonlinearly, which resulted in larger drag.
Bester and Behringer’s investigation into how the shape of a conical intruder falling into sand affects the forces it experiences is a beautiful example of how complex the interactions of everyday materials can be. According to their work, the aliens in our introduction should drop a pointy probe from very high up to make sure it gets buried — and also put some sensors on their probe to measure how its descent is interrupted by the force chains in the sand. The aliens may have imaginary science fiction technology that allows them to traverse light years, but even they may marvel at the countless collisions that affect the path of something they drop on the beach once they reach the Earth.
[1] The sum of the drag coefficient ($latex \int {h(z) dz}$) was calculated from the measured kinetic energy, and then the derivative of it was taken to obtain the drag coefficient h. Taking the derivative amplified the noise in the measurement. Bester and Behringer compared the sum $latex \int {h(z) dz}$. for different intruders to avoid this amplified noise.