It’s unusual to run a symposium as a PhD student, but anyone can do it! I was lucky to find a great mentor to guide me through the process. Together we organized 11 speakers, 2 workshops, and 11 poster presentations for a full day discussion on what soft matter, materials, and evolutionary biology have in common. From fire ants to spider silk, tooth enamel to lizard scales, and chemistry to computer science, there are lots of opportunities for soft-matter researchers to study biological questions.
The annual meeting of the Society for Integrative and Comparative Biology (SICB) is one of the core conferences for organismal biology. Originally called the “American Society of Zoologists,” the society changed its name to SICB in 1996 to emphasize the “integration” of different biological specializations. This commitment to interdisciplinary research made SICB the perfect home for my interest in biologically produced materials.
I’m interested in how biomaterials are created and diversify, a topic that draws on soft matter physics, mechanics, and evolutionary biology. There are a lot of exciting questions in this area, but because they are so interdisciplinary, there are not that many people who work on them. Interdisciplinary research often falls outside traditional departments and grant funding options, making these projects hard to design and run. They also require careful communication skills (if you talk to an engineer and an evolutionary biologist about the “evolution of a biomaterial” you might get two very different answers– the engineer might think of “material evolution” as a change during the material’s use (how does it respond to heat or light?), while the biologist might think about changes as the material developed with different organisms over millions of years). Nevertheless, I think interdisciplinary research questions are some of the most exciting and important, and luckily I’m not alone.
Together with my co-organizer, Dr. Mason Dean from the Biomaterials Department of the Max Planck Institute for Colloids and Interfaces, we organized the SICB symposium “Adaptation and Evolution of Biological Materials” (#AEBM #SICB2019) to highlight what is already being done in this field, and to encourage more biologists to start working with materials and soft matter.
Here are some highlights from our speakers:
Entanglement
Beyond “active matter” systems like fish schools or bird flocks, there are also collections of individual organisms that entangle together and behave like squishy, living materials. Prof. David Hu and Prof. Saad Bhamla presented on two different entangled soft matter systems: fire ant swarms and worm blobs. Both can act sometimes like a liquid and sometimes like a solid, depending on how the individuals link together. These systems can be described similarly to collections of molecules, complete with phase separation behavior!
These worm blobs are “active” and behave as viscoelastic fluids (both solid+liquid properties) — teaser video below. Come to the talk to learn more about these extreme biological system + active soft matter. https://t.co/iDYhPLkEri pic.twitter.com/MwlMcTdrmD
— Saad Bhamla (@BhamlaLab) January 2, 2019
Tunability
Unlike a lot of human-engineered systems, almost all biological materials have multiple functions. Dr. Beth Mortimer studies vibrational communication in spiders, worms, and elephants. Here she presented recent work suggesting the material vibration sensors built into spider legs might be tuned specifically for silk material properties — highlighting how silk has evolved to be both a structural and sensory material.
Assembly
Biological materials are famous for being made of simple, individual components that can assemble into complex structures on their own (i.e. “self assembly” without a human engineer). We had a lot of talks referencing this topic. Dr. Linnea Hesse studies the joints of branching plants to try and learn why they are so sturdy. She found that the vascular bundles that transport water (the equivalent of human capillaries for blood flow) adapt to external forces as the branch grows. This way the bottom of the branch is arranged differently than the top to optimize load bearing.
On a smaller scale, Prof. Matt Harrington presented on a new model of fiber formation from the velvet worm. Velvet worms shoot slime at their prey, which quickly hardens into fibers with strength comparable to nylon. If that wasn’t cool enough, these fibers can be dissolved in water and then later resolidify! Making them an intriguing model for new biodegradable plastics. Unlike spider silk, which is made of tiny highly ordered fibers, the “silk” of the velvet work seems to be made of relatively disordered charge-stabilized droplets.
Last but not least, Dr. Ainsley Seago has surveyed the colorful nanostructured scales of hundreds of species in two lineages of beetles. Her results suggest that even though these surfaces exhibit many different optical properties, they’re all likely assembled as liquids in a process remarkably similar to the assembly of cell membranes (called lyotropic assembly).
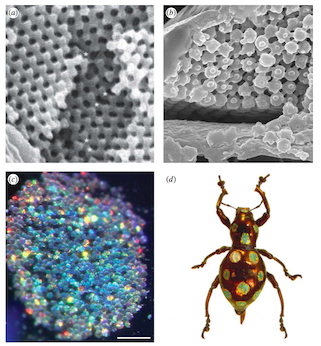
Image Analysis
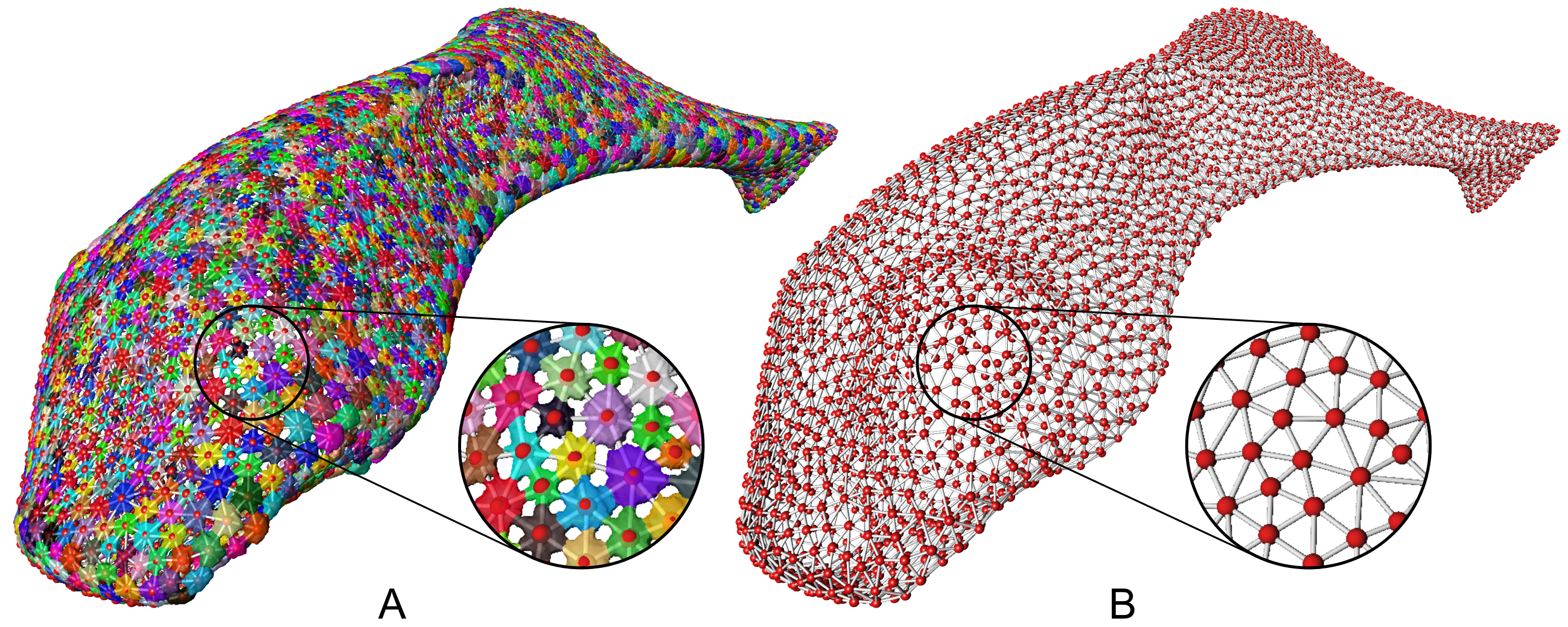
Dr. Daniel Baum is an expert on computational solutions for automated image analysis. He presented on common approaches for automatically selecting different parts of an image. This is really useful for studying material and biological systems with lots of similar repeating structures, and modeling how these systems respond to external forces. He presented examples of this work applied to the study of sharks and rays, whose soft cartilaginous skeletons are wrapped in a network of tiny, repeating, mineralized plates (called tesserae).
Hierarchy
The layered organization of materials at different scales (forming a hierarchy of structure) is important for many biological materials’ properties. Dr. Laura Bagge studies invisibility in deep sea ocean life, and she presented how the size of the tiny microfibrils that make up larger muscle fibers can change how opaque an organism is — larger microfibrils have fewer interfaces for light to interact with, allowing the whole body of some shrimp species to be transparent.
These kinds of hierarchies are more commonly associated with strength, as in the example that Dr. Adam van Casteren presented. He studies how enamel, the outer layer of the tooth, resists wear, showing work suggesting that different levels of the material structure (nanostructure vs. microstructure) might respond differently to evolutionary pressure. That means that these hierarchies might have evolved to protect against damage from different types of diets, i.e. abrasion from sand particles in plant-based diets versus fracture from breaking apart bones and shellfish.

Microfluidics
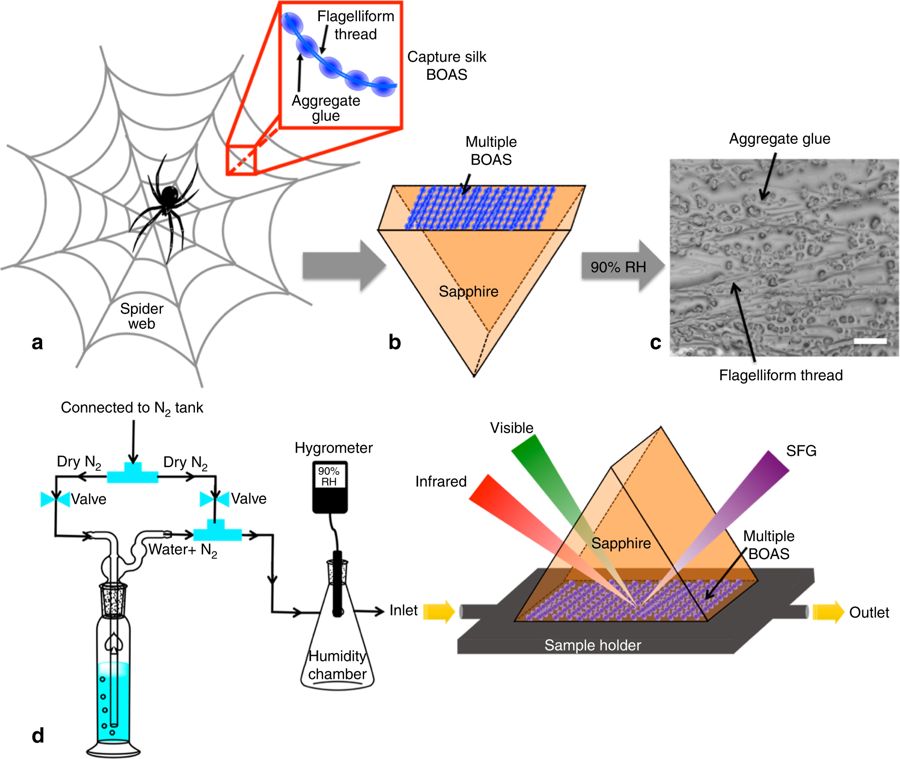
Fluid transport (both liquids and gases) is crucial for organism survival, so it’s no surprise that many biomaterials have been optimized for this function. Dr. Anna-Christin Joel presented work on how lizard scales and certain spider silks use capillary forces to manipulate fluids. The same capillary control has been harnessed to transport water droplets collected along the body to the mouth for drinking and to make capture silk stick more tightly to prey (by pulling waxes up from the surface of insects).
In a different application of fluid handling, Prof. Cassie Stoddard talked about the large eggs of emus. All eggs have pores that provide airflow to the growing chick, but the pores in emu eggs are forked not straight. This might help solve the challenge of getting enough breathable air into large eggs without weakening the shell enough that it could be crushed by the adult (interestingly this feature is also seen in dinosaur eggs!).