Original article: Emergent dynamics of laboratory insect swarms
Imagine yourself as a small fly called a midge (shown in Figure 1a). You used to live in a lake as a small larva with no concerns in life except swimming, eating, and growing. One day, you hid underwater and formed a cocoon around your body as it developed wings, legs, and antennae. A few days later, you swam to the surface and burst out of your cocoon as an adult fly — a male. As a new adult male, you find the clock ticking – you have only a few days to find a mate before you die.
Attracting a female is difficult for a tiny midge – how is she going to see you flying around? Fortunately, you can team up with hundreds of other male midges. Together, you fly above the lake in a stationary swarm that looks like a large cloud. Females can find this swarm and fly into it for their choice of mate.
In today’s study, Douglas Kelley and Nicholas Ouellette investigate how the motion of midges in a swarm helps the swarm stick together.
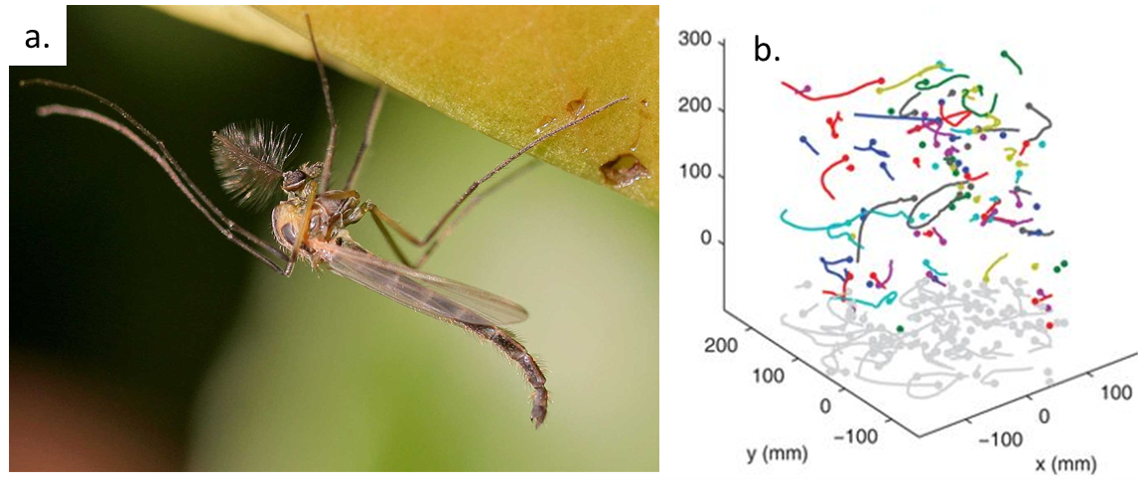
The researchers set up midge swarms in the laboratory and film them with three infrared cameras. They track all of the individual midges in the swarm and calculate their trajectories. Tracking dozens of small midges is not easy! First, they use the 2D images from the three cameras to locate the flies in 3D for each frame. Then, they use a technique originally developed for studying turbulent fluid flows to generate the trajectories over time. This technique uses the history of each midge to estimate where it is likely to be next and looks for midges in that area at the next time step. The resulting trajectories are shown in Figure 1b. The positions, velocities, and accelerations of the midges give clues about how the swarm moves.
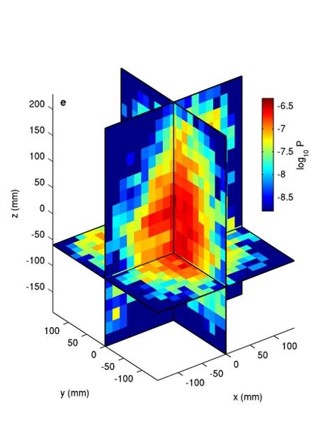
First, Kelley and Ouellette discuss the position of the midges. They plot the logarithm of the probability ($latex log_{10} P$) of finding a midge in a point in the swarm in three dimensions in Figure 2. Bright red represents a high probability of finding a midge, and blue represents a very low probability. Midge swarms are nearly symmetric, but larger swarms (of 100 individuals or more) are slightly taller than they are wide. This is unlike bird flocks, which are nearly two-dimensional.
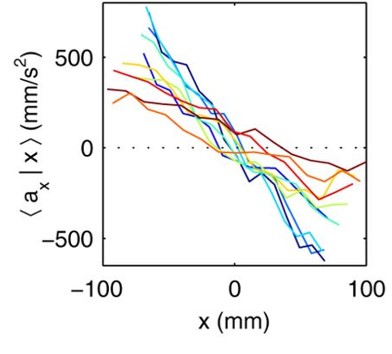
The researchers then investigate the velocities of the midges. Since the swarm is stationary, the average velocity of all the midges in the swarm is nearly zero. It turns out that the standard deviation, or the variation, of the velocities of the individual midges is more useful for understanding the motion inside the swarm. Midges fly twice as fast horizontally as vertically, just like birds in a flock. However, unlike flocks of birds, the midge swarms are not polarized — a midge does not tend to fly in the same direction as its neighbors. Finally, Kelley and Ouellette investigate the acceleration of the midges in the swarm. The midges are equally likely to turn in any direction, unlike birds or fish. Relative to their body size, larger animals have more inertia than smaller ones, and must exert a lot more effort to turn, accelerate, and decelerate. Thus, they tend to keep moving in the direction they are currently moving. Midges, on the other hand, can turn and easily move in any direction. Kelley and Ouellette find the average acceleration of the midges in the swarm in the x-direction as a function of the midge’s x-position, $latex \langle a_x|x \rangle$, in Figure 3. The midges tend to accelerate towards the center of the swarm, keeping the entire swarm together despite the midges’ constant motion. [1]
In this study, Kelley and Ouellette quantify a new type of swarming behavior. Unlike most other animal aggregations like bird flocks and fish schools, midge swarms stay in one place, helping female midges find the love of their very short lives.
[1] Surprisingly, when Kelley and Ouellette investigated the mean square displacement of the midges, they found that the midges act as if they are trapped in a box with walls.