Original paper: Structural predictor for nonlinear sheared dynamics in simple glass-forming liquids
We’ve all been there. We try pouring ketchup onto our fries from the bottle, but it doesn’t come out. So we tap the back of the bottle a few times, and suddenly, the ketchup rushes out and your entire meal is covered with it. Why does the ketchup exhibit such behavior?
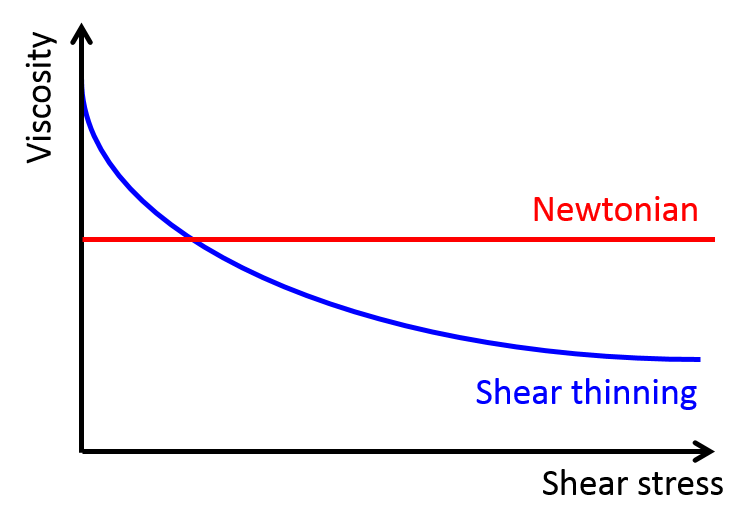
This behavior is called shear thinning, and only some special fluids exhibit it. For fluids, such as water and alcohol (these are called “classical” or “Newtonian” fluids) viscosity only depends on temperature. Therefore, if the temperature doesn’t change, the viscosity remains constant (see the red curve in Figure 1). However, in non-Newtonian fluids, viscosity depends on another variable called the shear stress. Shear stress is the stress felt by materials when they undergo deformation caused by slip or slide. In shear-thinning fluids, which are a type of non-Newtonian fluids, the viscosity decreases when the shear stress increases (see the blue curve in Figure 1). Ketchup, with other suspension fluids such as blood and nail polish, falls into this category of shear-thinning fluids. So, by tapping the ketchup bottle, we apply shear stress to the ketchup inside, causing the viscosity to drop and making the ketchup flow out of the bottle. But, even though this phenomenon has been on scientists’ radar for a long time, the microscopic mechanism for shear thinning is still unknown for certain fluids.
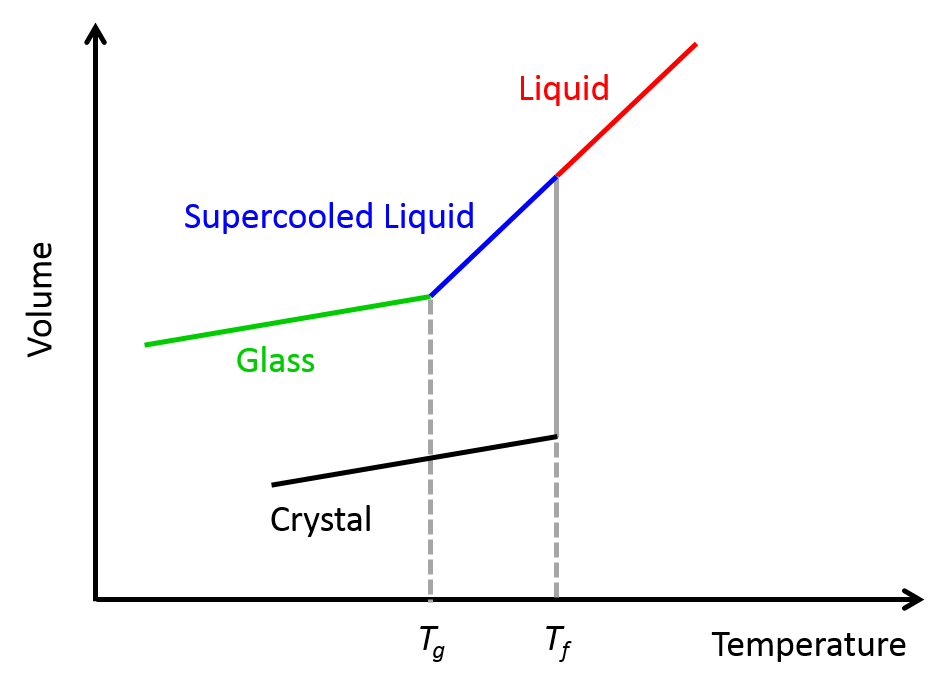
Another type of fluid that exhibits shear-thinning behavior is the “supercooled” liquids. As shown in Figure 2, when a liquid – any liquid – is rapidly cooled below its freezing point, instead of crystallizing and solidifying (like what we typically see when water freezes in an ice-cube tray), it forms a supercooled liquid. When the temperature of this highly viscous liquids drops even further below its glass-forming temperature, it turns into a disordered glass-like phase [1]. That is why supercooled liquids are also called glass-forming liquids.
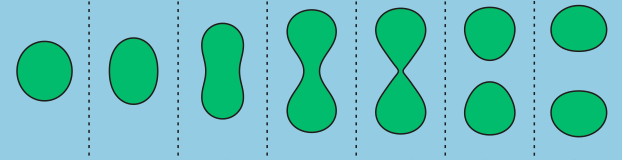
To understand the flow behavior of supercooled liquids, Trond Ingebrigtsen and Hajime Tanaka of the Institute of Industrial Science at the University of Tokyo ran molecular dynamics simulations. Molecular dynamics simulation is a computational method for studying the interactions of atoms or molecules. From the simulations, Ingebrigtsen and Tanaka were able to confirm what other scientists had previously suspected: shear thinning is linked to the increase in structural disorder of the liquid molecules (as illustrated in Figure 3(a) and 3(b)). To be more specific, it is linked to the structural disorder of molecules in the flow direction.
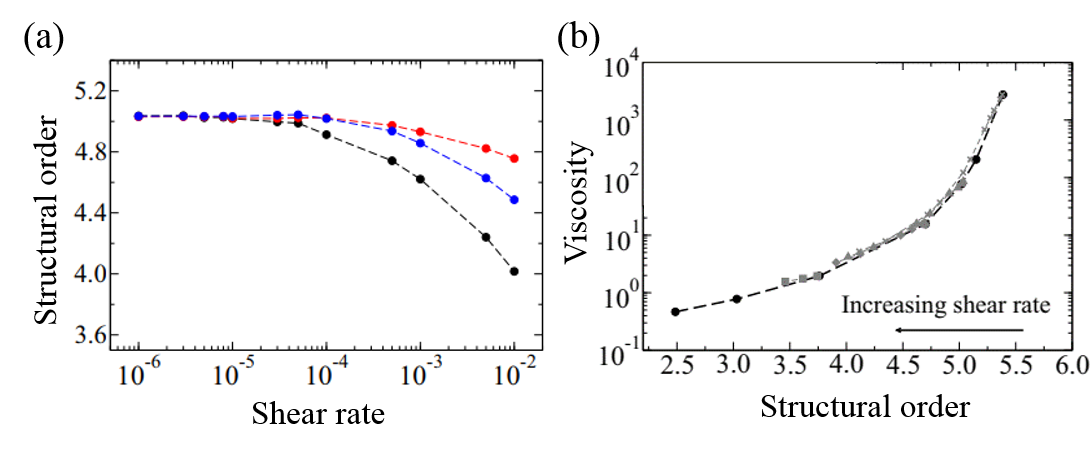
As a model for supercooled liquids, the authors chose to simulate a colloidal system, where molecules interact in a similar way to realistic fluids. After verifying that the simulates system acts like a supercooled liquid (for example, its viscosity decreases with increasing shear rate), they investigated the origin of shear thinning using this model. The molecular simulation revealed that as the shear rate increases, the molecular structure becomes more disordered. This is illustrated in Figure 3(a) and 3(b). More notably, the structural disorder was more prominent in the direction of the fluid flow compared to the structural disorder measured in any other directions relative to the flow. This can be seen from the black line of Figure 4(a), where the steep decrease of structural order could be observed with increasing shear rate.
Indeed, the structural disorder turned out to be the culprit behind the shear-thinning behavior in supercooled liquids. As shown in Figure 4(b), when the molecular structure becomes more disordered, the viscosity of the liquid decreases, a behavior expected in shear-thinning fluids. To understand this result, let’s picture molecules in the fluid. The shear applied in the direction of the flow would open up more space for molecules to rearrange themselves as the fluid expands, like it is shown in Figure 3(c). This leads to the decreased viscosity and the easier fluid flow.
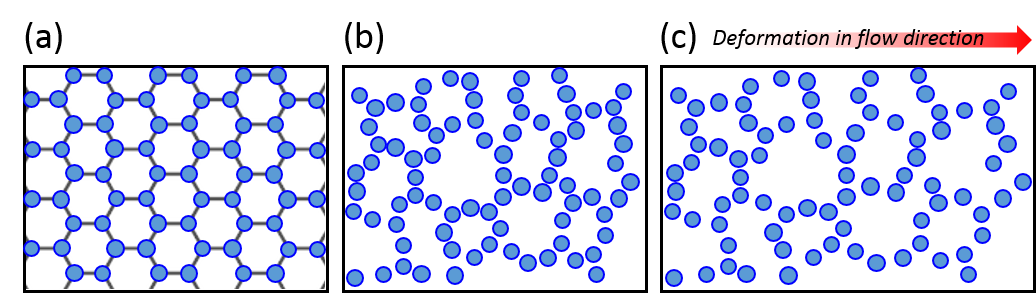
This study sheds light on the previously unknown mechanism of shear thinning in supercooled liquids. Ingebrigtsen and Tanaka, however, insert that the microscopic mechanism for their observation should be further studied to fully understand the shear-thinning behavior. So, next time a disaster happens on your fries, chill out and think that you are just carrying out a super cool non-newtonian experiment!
(This post was updated on March 4th, 2020 to answer a comment that was made on the French translation of this post.)
[1] Technically, glass isn’t a phase, though I used that word for simplicity. Glass is an amorphous solid that has a disordered molecular structure (unlike ice, which has a well-defined crystalline structure). See Figure 3(b) for a visualization of a disordered molecular structure.